在以往的研究中,人们将细胞按再生能力强弱分为不稳定细胞、稳定细胞与永久性细胞。由不稳定和稳定细胞构成的组织,为维持完整性,实质细胞丧失后会有同样表型的细胞再生取代。再生细胞来源于该组织完全未分化或部分分化的干细胞
[6]。例如,前列腺、乳腺的干细胞存在于腺体基底层或肌上皮层,表达CK5与P63的细胞群中
[7]。
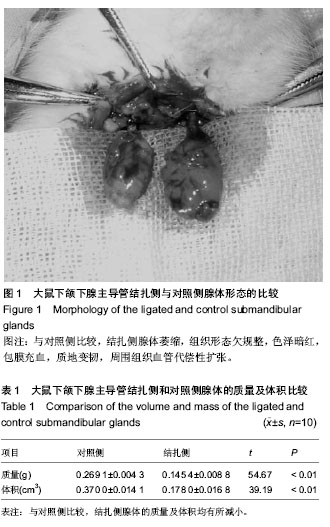
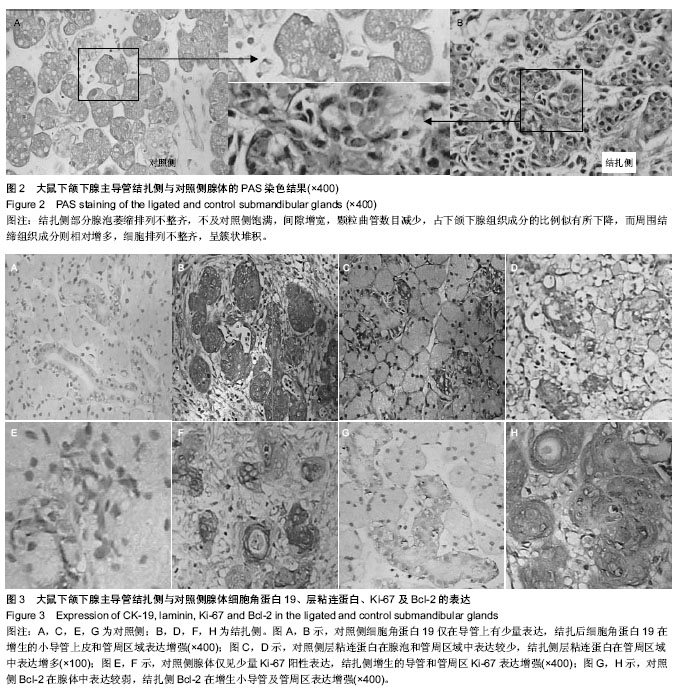
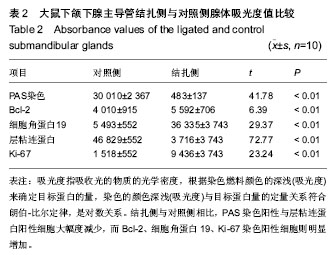
在动物实验中发现,将下颌下腺主导管结扎后,腺体组织的质量和体积均发生了减少,腺泡组织面积占腺体总面积的比例也发生减少,细胞分泌功能下降;而小导管结构大量增生,增生的小导管上皮细胞表达细胞角蛋白19阳性,在对照组腺体中,此区域表达细胞角蛋白19阳性的细胞非常少见。以往研究已证明,在与涎腺有同源性的肝脏损伤后的,在修复过程中,其卵圆形细胞(干/祖细胞)出现在肝门区,这些细胞表达细胞角蛋白19阳性,并能进一步分化为肝细胞和胆管细胞[8]。有学者在结扎鼠下颌下腺主导管后,分离到一群Sca-1+和c-Kit+的细胞,在体外培养中,保持了多角形,上皮样的形态结构,并具有增殖能力。在体外高浓度matrigel中培养可以分化为肝细胞,与GLP一同进行球形培养(spherical cultures),可以分化为胰腺细胞;而移植到鼠的肝脏中,能与受体肝小叶完全融合,它们在受体肝中表达白蛋白和α1-抗胰岛素,这群细胞还可分化为细胞角蛋白19阳性,导管样上皮细胞。说明这群细胞在体内体外均具有多向分化潜能,具有涎腺干细胞特征[9-10]。
细胞外基质是填充于细胞间的物质, 其成分除水、电解质,少量液相成分外,主要还有胶原、糖蛋白和蛋白多糖等。他们不仅是组织细胞结构的支持物,而且可以影响细胞的形态,调控细胞的正常代谢、迁移、增殖、分化以及信息传递,其中基底膜与细胞相互作用,调节细胞黏附、生长和分化的关系最为密切。层粘连蛋白是基底膜的特异的和主要的蛋白组成成分, 是一种由许多具有不同结构和功能区域构成的存在于基膜中的大分子非胶原性糖蛋白,在体内通过与其受体(如整合素类受体)结合,影响细胞的黏附、迁移、增殖、分化,从而发挥重要作用[11]。
在胚胎发育过程中,由非极性的间充质细胞转化为极性的上皮细胞,伴随有层粘连蛋白(基膜成分之一)的合成和分泌。细胞极性以其细胞膜表面出现两个不同的功能面即基底面和游离面为特征层粘连蛋白一经出现细胞即与基膜黏附[12],导致细胞表面和分子结构的不对称,也就具有了极性。有学者认为,基膜决定细胞的极性,层黏连蛋白的出现是细胞极性开始的基本成分。也有学者认为,上皮细胞仅在黏附于基膜时才能显现出极性在基膜形成之前表达于管周的细胞团,在成熟组织中多表达于基膜存在的部位。正常下颌下腺组织中,层粘连蛋白是腺泡及导管上皮基底膜的主要成分,在成熟下颌下腺导管细胞中,很少表达,而在小鼠的干/祖细胞中有表达[13]。
在本实验中,主导管结扎前,腺泡细胞表达层粘蛋白的阳性表达率,主导管结扎后,可见丛状层粘连蛋白阳性细胞主要出现在增生导管周围,也就是腺体发生再生反应的区域出现了层粘连蛋白胞浆阳性的细胞小集落,此区域细胞角蛋白19也存在阳性表达。有学者证明这些位于增生导管周围腺体发生再生反应区域的细胞,进行分离纯化后具仍有高黏附、高增殖等组织干细胞的相关特性[14-15]。
正常情况下组织中的成体干细胞通常处于静息状态,以对称分裂和非对称分裂两种方式生长,对称分裂形成两个相同的干细胞,非对称分裂生成一个分化细胞,而另一个仍作为干细胞保留下来,干细胞在整个增殖过程中处于相对静止。当受到损伤等情况时,干细胞的分裂方式会发生改变,处于相对活跃的增殖状态,以适应机体的需要。成体干细胞在组织结构中的位置相对固定,处于一个由干细胞、基底膜和调控细胞组成的微环境之中[16]。
Ki-67抗原是由相对分子质量为345 000和395 000的两条多肽链组成的核蛋白,作为一种核增殖抗原,其表达出现于除G0以外的G1,S,G2和M期的细胞周期中,M期达到最高。由于其在有丝分裂后迅速裂解或失去抗原决定簇,能较可靠地全面反映细胞群体的增殖活性,是判断细胞增殖活性的一种客观指标和特异性指标,其阳性率能反映细胞增殖情况,高阳性率意味细胞增殖迅速[17]。而正常涎腺细胞是一种终末分化的细胞,增值较慢[18]。
Bcl-2是细胞内的原癌基因,与细胞内的抗氧化损伤保护有关,其编码的Bcl-2蛋白是一种膜结合蛋白,定位于线粒体、内质网及核膜上,参与清除活性氧、阻抑膜脂质过氧化,与细胞内的抗损伤自稳态有关。Bcl-2可以表达在多种造血细胞、某些正常上皮(多为长寿命、具有增殖能力的外分泌导管上皮)、肠腺窝基底细胞等[19]。Bcl-2属于低丰度表达基因,仅在增生活跃的细胞有明显转录,在高度分化的正常组织中难以检测到。高表达Bcl-2可阻抑抑制各种因素包括物理、化学、生物等对细胞凋亡的诱导,延长细胞的存活时间[20]。
干细胞是一种尚未分化具有自我更新能力的细胞。在特定的条件下,成体干细胞可产生新的干细胞也能够按照一定程序分化形成新的功能细胞,从而使组织和器官保持生长和衰退的平衡。成体干细胞能维持于未分化状态不是由单一或者几个(oct3/4、LIF/SATA3等)因子决定的,是许多因子、信号通路共同作用的结果。Bcl-2在成体干细胞处于增殖状态时高表达,而在分化的细胞中表达很低。提示Bcl-2可能在维持成体干细胞自身稳定并处于未分化状态中起到了一定的作用。在实验组结扎后Ki-67和Bcl-2均高表达于增生导管周围也就是腺体发生再生反应的区域,提示此区域存在一群低分化的增殖活跃细胞[21]。
本实验发现,将下颌下腺主导管结扎后,腺体组织的质量和体积均发生了减少,腺泡组织面积占腺体总面积的比例也发生减少,细胞分泌功能下降;而小导管结构大量增生,增生的小导管上皮细胞表达细胞角蛋白19阳性,在对照组腺体中,此区域表达细胞角蛋白19阳性的细胞非常少见。主导管结扎后,腺体细胞对层粘蛋白的阳性表达升高,层粘连蛋白阳性细胞出现在增生导管周围。同时结扎后腺体表达Bcl-2和Ki-67均有增加,提示细胞增生活跃。作者认为在组织再生反应主要的发生部位既下颌下腺的导管和管周区域可能存在下颌下腺的成体干/祖细胞。