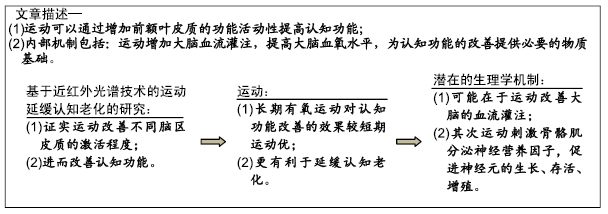
2.1 认知老化与大脑的皮质活动性相关 认知老化指随着年龄的增加,认知功能出现衰退的现象,其中认知加工速度和工作记忆对认知变量的影响最为显著。认知加工速度包括知觉速度、感觉速度;工作记忆是大脑对信息短暂性的加工和储存的能力[1],在日常生活中常表现为个体有想说的说不出的语言失败,通常称之为“舌尖现象”。认知老化具体表现为归纳推理能力、空间定向能力、数字能力、语义能力及词汇流畅能力的降低。当前对认知老化的理论学说包括:加工资源学说、共因假说、信息降格假说。加工资源学说即认知加工速度的增龄性改变,个体难以集中注意力对无关信息的抑制能力减弱。共因假说即影响感知觉的共同因素对大脑整体的信息加工产生广泛影响,伴随老化过程,大脑的神经信号表现为:模糊化、去独立性、去分化的特点。信息降格假说是指大脑将有限的资源进行感知觉的加工,资源不足时,导致认知任务表现差[2]。
认知老化过程中大脑具有补偿效应。认知老化与大脑皮质功能活动性的减少有关,且具有显著的增龄性特点[3]。认知功能更容易受到大脑整体的活动性减少[4],结构和功能减退的影响。影像学研究表明,在完成相同的认知任务的过程中年轻人的大脑皮质呈现出不对称的激活,在老年人中这种不对称性下降,此外老年人中大脑对相同的任务激活模式表现为后部到前部的转移,激活区域的转移,影像学研究表明在认知老化的过程中前额叶的激活程度及功能连接减弱。年轻人在完成情节记忆任务和言语工作记忆的过程中,主要激活了左前叶的皮质区,在提取任务阶段和空间工作记忆任务中,右前额皮质区域激活明显,对于特定任务的前额叶的激活程度具有明显的不对称性的特点,而在老年人中,这种不对称性显著下降。在执行任务的过程中,和海马区前部相比较,认知老化人群的内侧前额叶的功能连结减弱。这也进一步解释了老化造成大脑整体性的萎缩,使认知老化人群在执行特定认知过程中,大脑的激活程度和功能连结较年轻人差的原因。前额叶的结构和功能具有可塑性,通过有效的手段能够延缓前额叶的萎缩,改善前额叶的血流灌注,进而起到延缓认知老化的目的。
2.2 运动对不同脑区皮质活动性的影响
2.2.1 运动对大脑额叶皮质功能活动性的影响 基于近红外光谱的基础上运动对额叶皮质的研究主要集中在:前额叶皮质区域、额叶脑回、额回、眶回、中央前回、前扣带回、后扣带回;主要负责高级的认知功能:执行控制功能、注意功能、抑制功能、记忆功能、语言加工功能和情绪处理功能[4]。大脑前额叶皮质血液动力学变化常用的认知范式包括:stroop测试、flanker测试、GO/NO-GO测试、随机数字序列测试、修正后的sternberg测试、空间工作记忆测试,以此来评估短期工作记忆,大脑皮质的血液动力学应答[5-6]。
前额叶皮质对运动的应答程度受到运动类型的影响,在一过性的急性运动之后的认知测试时间在1- 5 min,运动强度过大,或是在低氧条件下运动,大脑的血氧含量通常较低,只有在中等强度的常氧环境下进行一过性的运动之后进行认知测试,大脑的激活程度才能处于最适水平,认知任务的表现处于最佳状态。此外,认知老化人群在急性运动之后前额叶激活的位置和年轻人相比具有差异性。研究表明,在年轻人中左前额叶背外侧区域和左前额极区的激活程度和反应时相关。而在老年人中,10 min中等强度的功率自行车运动,与右侧额极的激活程度相关[7]。额叶皮质体积的增加和平衡性表现相关,学习相关的大脑结构发生改变,这些结构的改变不仅仅限于皮质结构,同样包括杏仁核区域,而相关的功能链接减少的区域包括神经基底节,尤其是尾状核区域。
在运动条件下脑血氧的研究大部分集中在前额区域,这可能与运动后排汗和头发的厚度有关,在这样的条件下,容易造成以偏概全的结论。因此在今后的研究中,针对不同个体,通过检测不同脑区的血氧含量,间接判断脑区是否被激活及其激活程度,通过筛选有效的干预方案最终实现目标脑区的能力增强。近来的研究也强调,局部的血氧含量不能代表整体,应该加强多指标相结合、整体指标和局部指标的结合、中枢神经系统和外周神经系统的结合、氧合血红蛋白和脱氧血红蛋白的同时报告及不同区域的指标相结合的方法,提高参数的可靠性和有效性。此外应该充分考虑运动条件,被试的身体状况(心肺疾病和心血管疾病)都是影响运动介导的大脑血氧含量的中介变量。
2.2.2 运动对运动皮质功能活动性的影响 运动皮质位于额叶皮质的后部,主要包括运动皮质/前运动皮质和辅助运动皮质,主要负责计划、自动控制、自主运动过程中的学习和执行。
大强度运动导致的运动皮质血氧含量的下降是诱发运动疲劳的可能性原因。次最大强度的运动中,大脑运动皮质血氧含量显著下降。力竭运动不仅会造成运动皮质血氧含量的下降,同时前额叶皮质血氧含量也呈现出降低的趋势。这也进一步验证了,力竭运动不仅会造成肌肉等的周围系统的疲劳,同样会造成中枢疲劳,大脑皮质血氧动力学工作效率的下降伴随着脑功能的下降,运动神经元对工作肌肉的控制能力的降低,这也是力竭运动过程中认知能力下降的原因。
基于近红外光谱基础上,大脑血液动力学的特点是在被长期调控下不断适应的结果,并随着运动技能学习的变化而变化。大脑运动皮质与动作学习和技能获得相关,研究表明在专业运动员中,对刻意动作的反复练习,这些程序性的技能学习,运动皮质血液动力学的加工过程是在间接演化过程中被最优化的结果。运动皮质的变化与动作学习(手眼协调的动作练习中,以及多关节精细动作这样的任务中)过程中,大脑皮质的血氧动力学变化相一致 [8-12]。
2.2.3 运动对颞叶皮质功能活动性的影响 颞叶主要包括颞回、梭状回、主要的听觉皮质和海马回,海马区主要负责记忆、视觉、听觉、语言、情绪的加工过程。
有氧能力和颞叶皮质的激活程度呈正相关。在中老年人中,认知表现随着年龄的增加逐渐下降,有氧运动能力的提高能够改善认知表现,这些改善的认知表现包括计划、抑制功能和工作记忆功能、执行功能,这些认知功能和颞叶激活程度的增强有间接的关系。身体活动可以增强老年人的有氧能力,在老年人中,有氧能力越强,海马区的体积、大脑区域的活动、执行功能、空间记忆能力较高。研究表明,人体的有氧能力改善,能够提高大脑中轴线上皮质区域的功能连接及威斯康汀的卡片分类测试Wisconsin card sorting task 的任务表现。功能磁共振研究表明4个月的有氧运动不仅使老年人海马区的血流量增加,在利用全脑网络连接联结分析发现了海马和前扣带回联接的增强,伴随认知功能提 高[13]。因此有氧能力、大脑皮质的激活程度以及认知功能三者之间呈现出积极的正相关,且颞叶皮质区域额激活程度是有氧体适能水平和执行功能之间的重要中介变量[11,14]。
2.3 大脑皮质的激活程度和运动负荷相关 运动负荷不断增加,大脑皮质的区域活动增加。在运动和认知任务的过程中,影像学研究表明大脑皮质血氧水平增加是大脑血流增加的标志,伴随神经代谢激活,在此过程中,氧合血红蛋白的含量与氧气的运输、提取,以及整体血容量、血氧含量相关。在运动负荷不断增加的过程中,大脑通过更高水平的自动调控即更为集中的注意、协作、反应和自动控制等,完成复杂的工作任务。当运动强度增加到一定程度时,大脑为了平衡资源配置,相关的神经活动受到抑制,大脑的激活程度下降[15]。运动通过提高大脑皮质活动性改善认知功能。横断面研究表明运动可以改善特定的认知功能,例如:有氧运动可以改善背外侧前额叶皮质的活动性,而这些区域与空间工作记忆和抑制功能相关。而电生理的研究进一步证实了运动增加前额叶皮质的功能活动且改善特定的认知表现。ERICKSION等[11]的研究表明,对老年人进行中低强度的健步走或者健身操运动可以增大背外侧前额叶灰质的体积、改变前额叶皮质和皮下的功能激活,VOELCKER 等[16]的研究对老年人进行1年的有氧训练,发现这些运动组老年人抑制任务中额中回、额下回的激活减少,丘脑和纹状体的激活增加。其次,随机对照研究表明,即使10 min的急性运动也可以改善认知老化人群的认知表现,且这种认知表现和左侧额极激活水平相一致。
运动对大脑血氧带来整体的积极效益,使静息状态下的大脑血氧升高;但运动负荷过大所导致的运动疲劳,也会产生许多负面效益。长时间的运动,所导致的能量供给无法满足实际需要,引发的机体的中枢疲劳及周围疲劳,最终会影响中枢的有效运转[6]。在亚极量的运动过程中会导致身体同侧运动皮质和身体对侧运动皮质血氧水平的下降,大强度运动诱发的运动疲劳导致前额叶运动皮质、次级运动区皮质血氧含量下降,这进一步验证了运动疲劳诱导的血氧水平下降,不仅发生在周围骨骼肌,同样发生在中枢神经系统,诱发皮质血氧动力学的变化。在力竭运动过程中,大脑血氧水平的下降是大脑功能下降的标志,运动神经元发送的支配骨骼肌运动的信号减弱,这也就进一步解释了高强度运动过程中认知能力下降的原因[17-18]。
长期的有氧运动改善认知功能。一次性的有氧运动(≤20 min)可以改善视觉和空间感觉、短期记忆能力,但是这种作用却是即刻的。长期适宜的有氧运动(≥3个月),均有改善记忆和执行功能的作用。长期的运动,每次运动持续时间大于40 min,持续时间大于6个月的中低强度的运动无论对短期记忆和长期记忆均有改善的效果。COLCOMBE等[19]也进一步验证了3个月的有氧中等强度的运动可改善言语学习和记忆、增进心肺健康并增加大脑的体积。Meta分析表明长期的中等强度运动能够对记忆起到改善作用,且效果显著,持续时间较长[9]。有趣的是,BROWN等[20]进一步通过动物实验验证了这种区域性的大脑体积的特殊改变是大脑神经形成的标志。提示:长期运动改善认知功能,这可能与运动改善大脑血氧含量,进而起到改善这些脑区认知的作用相关[5]。而这些结果不一致,可能与运动强度、频率、持续时间、性别和一些参与人群多寡所导致的中介变量干扰所致,进一步的研究需要控制相关因素。而上述研究即使对于相同结果,在分析讨论的解释方面也存在差异,可能需要进一步结合更加严谨的任务态的近红外光谱和一致的量表,在严格控制中介变量的情况下,验证认知老化的认知功能的改变与哪些脑区有关。从上述研究中也可以看出,认知老化人群存在多个脑区的血氧含量值的下降,且主要存在于认知(记忆和执行)等脑区,这些脑区血红蛋白值的改变可能与记忆执行功能的下降相关,但目前还不十分清楚血氧含量的下降是出现记忆和执行功能下降的原因还是结果。运动和认知之间具有复杂的相关关系,无论什么形式的有氧运动在改善特定记忆功能方面存在异质性。换言之,并不是所有的有氧运动都可以改善认知功能并起到增强记忆的作用,最有效的改善特定记忆的有氧运动形式,取决于该种干预方式对个体是否最适宜,同时也要考虑到年龄、心血管的健康程度等也是重要的中介变量。
总体而言,有氧运动结合力量训练、柔韧性练习的效果比单一一种形式对认知的改善作用更好,这可能是由于不同的训练方案的结合会动员到更多的脑区,可以更好的减少与增龄性相关的心肺和骨骼肌下降及认知功能下降[17]。
2.4 运动改善认知老化的可能性机制 运动改善认知老化的可能性机制归结起来包括:神经递质学说、血管功能学说、中介变量学说。神经递质学说是指:运动通过大脑血流增加,大脑前额叶激活程度明显,同时伴随着大脑营养物质的增加和代谢的增强,神经递质释放,大脑的抗炎症水平和抗氧化能力提高,大脑的内稳态和平衡的微环境得到维持,从而促进神经元的存活、增殖、分化、迁移[21]。血管功能学说是指运动负荷刺激心肺功能,改善血管内皮功能,提高大脑的血流灌注,进而起到提高认知的作用。中介变量学说是指不同运动项目本身作为新异刺激,提高大脑的功能活动性,同时集体运动项目,人与人相互合作和促进人际交往的功能,有利于各种认知过程的重复使用,改善脑网络的工作效率,起到延缓认知老化的作用[22]。当前从理论上解释运动改善认知老化的内在机制,可以认为体育运动通过多路径对认知老化产生作用,各路径之间彼此独立或者相互影响,有待进一步证明。因此下一步的研究,应从不同路径之间探讨运动改善认知老化的可能性机制,以体育学为依托,借助心理学、神经科学、教育学等多学科交叉融合为运动改善认知老化互补的、全方位的证据。
2.4.1 运动通过改善大脑微血管的血流灌注提高认知功能[23] 大脑微血管的血流灌注受到血管功能的影响。血管风险因素包括:糖尿病、高血压、高脂血症,高脂血症对整体的健康和认知功能的损害作用尤为明显,对终末器官和脉管系统产生广泛影响。血管风险因素所导致的后果由最初细微的大脑结构改变,认知功能的减退,逐渐发展成为痴呆。同样这种逐渐积累的微血管的变化,血管周脂质变性,与大脑的白质相关,尽管白质的代谢需求比较低,但和大脑灰质相比,大脑的白质更容易受到缺血的伤害,白质脑血管的血流灌注减少,增龄性相关的抗氧化能力减弱。尽管白质易受缺血的影响已经得到普遍证明,但是从轻微的认知障碍到中度认知障碍大脑白质变化的相关研究相对较少,因此大脑认知功能减退发病前伴随着大脑结构整体性的变化是认知功能减退和痴呆的早期信号[10,24]。
运动通过提高血管内皮功能,改善大脑微血管的血流灌注。大脑微血管的血流灌注受到整体血流的影响,运动可以通过增强血管功能,改善前额叶脑区的血流量,增强大脑的可塑性。这可能与运动改善了血管内皮功能相关,内皮祖细胞是血管内皮细胞的前体细胞,具有促进血管内皮新生的作用[25]。研究表明内皮祖细胞对脑损伤小鼠血管具有修复作用[26],运动可以促进内皮祖细胞的增殖和迁移,进而起到改善血管内皮功能的作用[27]。
2.4.2 脑源性神经营养因子是介导运动改善记忆功能的生理学基础 脑源性神经营养因子是一种神经营养因子,可以通过相关调节通路刺激神经皮质,诱发神经元结构的改变,参与神经修复过程,是介导运动与大脑认知方面重要的媒介。脑源性神经营养因子水平在30-39岁达到顶峰,之后逐渐下降,研究表明,无论是在运动组还是在久坐不动的对照组,脑源性神经营养因子水平在年轻人中几乎是老年人的2倍。脑源性神经营养因子可以在周围组织中被合成,这些组织包括骨骼肌、肝脏、脂肪组织、内皮细胞和免疫细胞,在通常情况下,大脑的合成占到75%。循环血清中的脑源性神经营养因子可以透过血脑屏障,参与神经细胞的生长、存活及增殖[28]。
脑源性神经营养因子在促进突触重塑和形态学的复杂性过程中起到重要作用,首先,其可以通过增加啮齿类动物和中青年人的静息状态下海马区的灌注及增加齿状回、内嗅皮质中树突的复杂性和数量,从而间接促进神经元的形成。在脑源性神经营养因子敲除的大鼠中,颗粒下层的神经干细胞显著减少,增殖中的神经干细胞呈现出死亡的状态,认知能力严重下降[29]。脑源性神经营养因子的表达可以显著促进神经干细胞的分化转移和成熟,很大程度上决定神经干细胞的分化,调控神经加工的过程,突触可塑性。其次,神经营养因子也可通过修复大脑血管内皮生长因子介导的突触可塑性,有效改善大脑的结构和功能。敲除脑源性神经营养因子基因大鼠由于神经可塑性降低,出现大脑海马区体积的减少和认知能力的下降等症状,即使通过运动也不能逆转神经可塑性水平的降低,以及大脑海马区体积的减少和认知能力的下降[30]。运动不仅使细胞的存活增加,还可以使海马区和皮质神经元数量上升,人体试验研究表明,有氧运动促进海马区脑源性神经营养因子的表达是通过促进海马区的脑源性神经营养因子基因表达得以实现的[31]。且人体大脑中脑源性神经营养因子的表达对有氧运动强度呈现出一种剂量反应。
这说明,脑源性神经营养因子可以促进神经元的形成,促使新生神经元进入循环,为海马区神经形成提供稳定的物质基础;也是血管生成、大脑海马区结构和功能重塑的基础。神经营养因子障碍可引起整个机体代谢和免疫功能紊乱,导致海马区萎缩,痴呆、抑郁等精神类疾病发生,因此脑源性神经营养因子是大脑内稳态的内置调节机制。海马区神经营养因子的相关调控通路见表1。

2.4.3 运动诱导组织蛋白酶B可以促进记忆功能的改善 动物实验研究表明,组织蛋白酶B可以增加成年大鼠海马区的神经形成及其空间记忆功能。组织蛋白酶B敲除的大鼠中,空间记忆功能能力下降。人体试验研究表明,跑台运动可以增加循环血液中组织蛋白酶B的水平,成年人海马区的神经形成,齿状回颗粒细胞的水平以及海马区蛋白11的水平降低。跑步运动能够改善记忆功能以及成年人的神经形成。组织蛋白酶B水平与健康程度以及海马区水平上的记忆功能相关。在成年人海马区的祖细胞中,组织蛋白酶B增加了神经营养因子的水平[32]。体内实验表明,运动增加了血浆中组织蛋白酶B的水平[33],同时海马区组织蛋白酶B基因表达水平也增加,表明了组织蛋白酶B对大脑直接和间接的作用[34]。
组织蛋白酶B能够促进海马区神经营养因子(脑源性神经营养因子)的表达,间接的促进神经形成[35]。研究表明,4个月跑步运动能够使骨骼肌组织蛋白酶B基因的表达增加,同时伴随着血浆中组织蛋白酶B蛋白水平的提高。此外,动物实验研究表明,组织蛋白酶B能够穿过血脑屏障,尽管组织蛋白酶B不能直接影响海马区细胞的增殖,但是其能够促进脑源性神经营养因子小RNA表达的增加,还可以增加脑源性神经营养因子蛋白的表达,以及促进双皮质素水平的增加,是一种可以促进神经迁移以及具有神经保护作用的蛋白。此外,组织蛋白酶B直接参与运动诱导海马区神经形成和海马区功能改善。MOON等[34]通过敲除大鼠中组织蛋白酶B基因,野生型大鼠运动可以诱导空间记忆功能的改善,但是缺少组织蛋白酶B基因之后,在实验室条件下,自主游泳运动不能够诱导同样的作用。也就是说,组织蛋白酶B在一定条件下可以介导运动改善海马区的神经形成、记忆和学习功能[36]。但是组织蛋白酶B从多大程度上决定运动诱导认知功能的改善有待进一步研究。
在此基础上,研究发现通过与拉伸运动的对照组相比,抗阻运动在改善推理能力方面更佳,抗阻运动同样可以促进海马区细胞的增殖,改善空间认知能 力[9],这也就说明了抗阻运动对于改善认知功能有不同的作用,也许在特定的执行功能方面效果更好。认知能力除了受到运动类型的影响外,运动应答的强弱也会受到个体差异的影响,基于此推测是由于基因的变化的影响的结果[14]。
2.4.4 有氧运动和抗阻运动均可以诱导的整体的纤连蛋白结构域包含蛋白5(fibronectin type
domain-containing protein 5,FNDC5)水平增加 FNDC5穿过血脑屏障,到达中枢神经系统,诱导海马区脑源性神经营养因子的表达,促进相关的神经形成认知功能的提高。人体研究表明,无论是急性还是长期的有氧和抗阻运动都能够诱导FNDC5表达的提高,运动刺激了肌肉和大脑的内分泌回路,通过PGC-1α和FNDC5,增加了脑源性神经营养因子的表达,最新的动物实验研究也进一步验证了在阿尔兹海默大鼠中,FNDC5和鸢尾属在调节突触可塑性以及记忆功能的作用[37]。

2.4.3 运动诱导组织蛋白酶B可以促进记忆功能的改善 动物实验研究表明,组织蛋白酶B可以增加成年大鼠海马区的神经形成及其空间记忆功能。组织蛋白酶B敲除的大鼠中,空间记忆功能能力下降。人体试验研究表明,跑台运动可以增加循环血液中组织蛋白酶B的水平,成年人海马区的神经形成,齿状回颗粒细胞的水平以及海马区蛋白11的水平降低。跑步运动能够改善记忆功能以及成年人的神经形成。组织蛋白酶B水平与健康程度以及海马区水平上的记忆功能相关。在成年人海马区的祖细胞中,组织蛋白酶B增加了神经营养因子的水平[32]。体内实验表明,运动增加了血浆中组织蛋白酶B的水平[33],同时海马区组织蛋白酶B基因表达水平也增加,表明了组织蛋白酶B对大脑直接和间接的作用[34]。
组织蛋白酶B能够促进海马区神经营养因子(脑源性神经营养因子)的表达,间接的促进神经形成[35]。研究表明,4个月跑步运动能够使骨骼肌组织蛋白酶B基因的表达增加,同时伴随着血浆中组织蛋白酶B蛋白水平的提高。此外,动物实验研究表明,组织蛋白酶B能够穿过血脑屏障,尽管组织蛋白酶B不能直接影响海马区细胞的增殖,但是其能够促进脑源性神经营养因子小RNA表达的增加,还可以增加脑源性神经营养因子蛋白的表达,以及促进双皮质素水平的增加,是一种可以促进神经迁移以及具有神经保护作用的蛋白。此外,组织蛋白酶B直接参与运动诱导海马区神经形成和海马区功能改善。MOON等[34]通过敲除大鼠中组织蛋白酶B基因,野生型大鼠运动可以诱导空间记忆功能的改善,但是缺少组织蛋白酶B基因之后,在实验室条件下,自主游泳运动不能够诱导同样的作用。也就是说,组织蛋白酶B在一定条件下可以介导运动改善海马区的神经形成、记忆和学习功能[36]。但是组织蛋白酶B从多大程度上决定运动诱导认知功能的改善有待进一步研究。
在此基础上,研究发现通过与拉伸运动的对照组相比,抗阻运动在改善推理能力方面更佳,抗阻运动同样可以促进海马区细胞的增殖,改善空间认知能 力[9],这也就说明了抗阻运动对于改善认知功能有不同的作用,也许在特定的执行功能方面效果更好。认知能力除了受到运动类型的影响外,运动应答的强弱也会受到个体差异的影响,基于此推测是由于基因的变化的影响的结果[14]。
2.4.4 有氧运动和抗阻运动均可以诱导的整体的纤连蛋白结构域包含蛋白5(fibronectin type
domain-containing protein 5,FNDC5)水平增加 FNDC5穿过血脑屏障,到达中枢神经系统,诱导海马区脑源性神经营养因子的表达,促进相关的神经形成认知功能的提高。人体研究表明,无论是急性还是长期的有氧和抗阻运动都能够诱导FNDC5表达的提高,运动刺激了肌肉和大脑的内分泌回路,通过PGC-1α和FNDC5,增加了脑源性神经营养因子的表达,最新的动物实验研究也进一步验证了在阿尔兹海默大鼠中,FNDC5和鸢尾属在调节突触可塑性以及记忆功能的作用[37]。