Tet-off/Tet-on基因表达系统是Gossen和Bujard等构建的,它可以有效调控基因表达的时间和水平
[10]。pTet-on质粒是转染真核细胞后表达RTTA,RTTA必须与四环素衍生物强力霉素结合后才能成为顺式作用元件,并作用于反应质粒pTRE-2Hyg上的TRE,从而激活转录,启动下游的目的基因的表达。TRE具有开关功能严密、特异性强、诱导效率高、基因表达水平高、对细胞无毒性等特点。该系统不仅能提供高水平的表达,而且可以实现剂量依赖调控
[11]。该表达体系包含2 个关键质粒:一个是表达转录调控蛋白的质粒(pTet-on);另一个是携带并表达外源基因的质粒(pTRE-2hyg),即当加入强力霉素时外源基因表达,去除强力霉素时外源基因的表达关闭,而且表达水平随四环素浓度的增加而增加。

PTHrP是一种具有复杂结构和功能的激素原,它广泛分布于机体的多个组织器官,通过自分泌、旁分泌以及胞分泌的方式发挥不同的生理作用。PTHrP的结构非常复杂,可以被激素原转化酶家族加工为至少3个功能片断,即氨基酸末端的PTHrP(1-36),中间片断的PTHrP(38-94),以及羧基端的PTHrP(107-139)。研究表明PTHrP(1-36)是PTHrP的保守分泌形式,可以促进肾脏钙的重吸收和延缓骨质疏松的发生
[6],PTHrP (38-94)可以通过激活IP(3)-DAG-PKC通路来维持局部钙离子浓度,钙离子的浓度又对成骨分化起着重要的作用
[7]。其中羧基端的PTHrP(107-139)具有抑制破骨细胞活性、促进成骨细胞增殖和促进成骨的特性,又被成为骨抑制素
[6,8],但它在骨骺干细胞和软骨细胞的生长和分化中是否有作用、有何作用尚不清楚。
增殖细胞核抗原是一种反应细胞增殖状况的指标
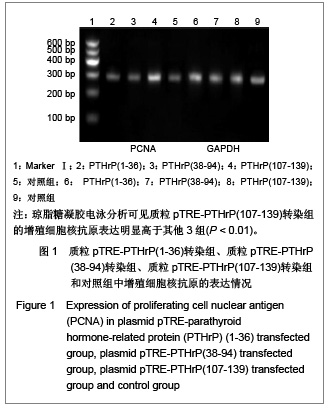
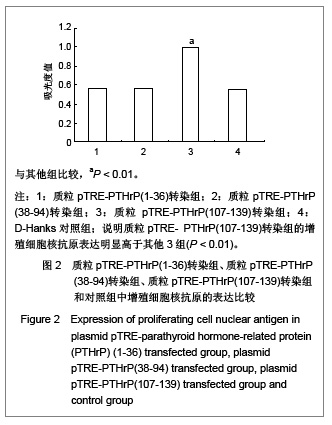
[12],实验中观察到随着加入强力霉素后,质粒pTRE-PTHrP (107-139)转染组的增殖细胞核抗原表达明显高于质粒pTRE-PTHrP(1-36)转染组、pTRE-PTHrP(38-94)转染组和对照组,证实了PTHrP(107-139)有促进细胞增殖的作用,而质粒pTRE-PTHrP(1-36)转染组、pTRE-PTHrP (38-94)转染组和对照组之间增殖细胞核抗原的表达没有显著差异,说明PTHrP(1-36)和PTHrP(38-94)对细胞的增殖能力没有明显影响。
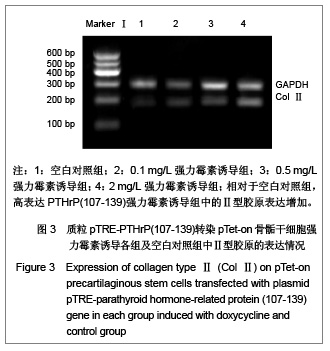
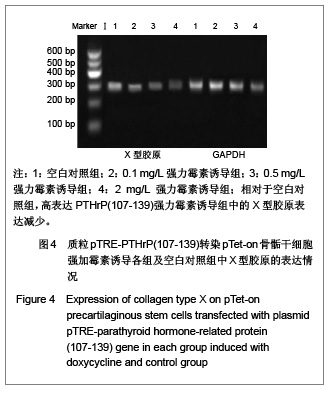
Ⅱ型胶原是骨骺细胞增殖时的特异性增殖基因,也是骨骺干细胞的标志蛋白之一
[13],而X型胶原的高表达则一般认为是肥大软骨细胞的标志
[14]。在实验中,相对于空白对照组,高表达PTHrP(107-139)诱导组中的Ⅱ型胶原表达增加,而X 型胶原表达减少,说明PTHrP (107-139)有抑制软骨细胞分化的作用。
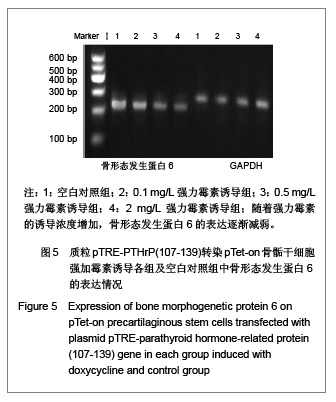
骨形态发生蛋白有诱导软骨细胞发生和促使软骨细胞终末分化的作用。有研究发现骨形态发生蛋白6参与了骨骺生长板中PTHrp-Ihh信号轴的负反馈调控,骨形态发生蛋白6可促使骨骺区细胞成熟,并上调的Ihh表达,而PTHrP抑制软骨细胞成熟是间接通过抑制骨形态发生蛋白6的表达来完成的
[2]。实验中也观察到随着强力霉素的诱导浓度增加,即PTHrP(107-139)表达的增加,骨形态发生蛋白6的表达逐渐降低。
Ihh通过PTHrP信号来调控软骨细胞分化的速率和促进软骨细胞增值
[1]。Ptc是其受体,在骺板中广泛分布[15]。当Ihh与Ptc结合后,激活Ptc等目的基因的转录调控,促进了关节周围软骨膜中PTHrP的分泌,而PTHrP在前肥大软骨期的大量表达又会抑制Ihh的分泌,即PTHrp-Ihh负反馈信号轴(也被称为负反馈信号环)
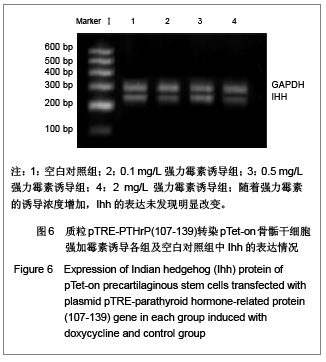
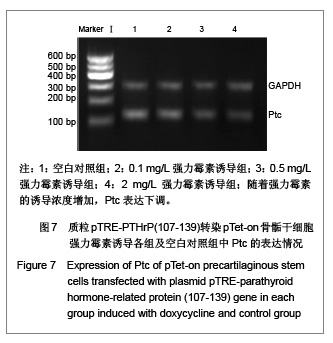
[1]。在0.5 mg/L、2 mg/L强力霉素诱导组中,Ptc表达下调,可能是因为一种负反馈调节机制。而Ihh的表达未发现明显改变,这与以往报道的PTHrP表达增加会抑制Ihh的表达(即负反馈调节)这一结论不符,可能是因为实验方法的影响,也可能是PTHrP(107-139)与完整PTHrP区别点所在,这有待于进一步的研究去证实。
Sox基因是编码转录因子的高迁移组盒子超家族中的一员
[5],其中Sox9在软骨发育、成熟过程中对维持细胞的表型及增强Ⅱ型胶原基因表达和促进Ⅱ型胶原合成增加方面发挥重要作用。有研究表明PTHrP通过Sox9来抑制软骨前体细胞向肥大软骨细胞转化
[4]。在实验中,在PTHrP(107-139)高表达组中(即2mg/L 强力霉素组),Sox9的表达也明显增强。
从以上研究结果可以看出:质粒pTRE-PTHrP(107-139)转染组骨骺干细胞的增殖细胞核抗原、Ⅱ型胶原表达明显增加,而X型胶原表达减少,说明PTHrP (107-139) 在促进骨骺干细胞增殖、抑制骨骺干细胞分化中发挥着重要作用。而质粒pTRE-PTHrP(1-36)、质粒pTRE-PTHrP(38-94)转染组骨骺干细胞的增殖细胞核抗原表达与对照组相比差异无显著性意义,说明亚基因PTHrP(1-36)、PTHrP(38-94)对PCSCs的增殖并无明显作用。在强力霉素诱导的质粒pTRE-PTHrP(107-139)转染组中Sox-9表达明显增强,Ihh表达未见明显变化,而 Ptc、骨形态发生蛋白6表达明显降低。说明THrP(107-139) 可能是通过调控Sox-9、Ptc、骨形态发生蛋白6的表达来促进骨骺干细胞增殖并抑制其分化的。而且上述因子的表达量与强力霉素存在剂量依赖性,再次表明了pTet-on质粒系统在PCSCs能有效的调控外来基因的表达。
由于多质粒的稳定转染成功率比较低,实验中尝试瞬时转染的方法,初步探讨了PTHrP亚基因在骨骺干细胞的增殖和分化中所发挥的作用,也揭示了PTHrP(107-139)与其他相关因子调控关系,对于PTHrP(107-139)表达增加未能使Ihh的表达发生明显改变这一点,还有待于进一步实验研究。
基金资助:中央高校基本科研业务费专项资金资助项目(4101052)。
作者贡献:设计由第一、六作者完成,实施由第一、二、三、四、五作者完成,评估由第一、六作者完成,均经过系统培训。
利益冲突:课题未涉及任何厂家及相关雇主或其他经济组织直接或间接的经济或利益的赞助。
伦理要求:未涉及与相关伦理道德冲突的内容。
作者声明:文章为原创作品,数据准确,内容不涉及泄密,无一稿两投,无抄袭,无内容剽窃,无作者署名争议,无与他人课题以及专利技术的争执,内容真实,文责自负。