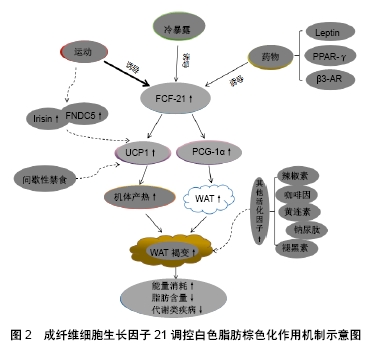
[1] KLIEWER SA, MANGELSDORF DJ. A Dozen Years of Discovery: Insights into the Physiology and Pharmacology of FGF21.Cell Metab. 2019;29(2):246-253.
[2] RODRÍGUEZ A, BECERRIL S, EZQUERRO S, et al. Crosstalk between adipokines and myokines in fat browning.Acta Physiol.2017; 219(2):362-381.
[3] LEE P, SWARBRICK M, GREENFIELD J. The Sum of All Browning in FGF21 Therapeutics.Cell Metab.2015; 21(6):795-796.
[4] FISHER FM, KLEINER S, DOURIS N, et al. FGF21 regulates PGC-1α and browning of white adipose tissues in adaptive thermogenesis. Genes Dev.2012;26(3):271-281.
[5] HONDARES E, IGLESIAS R, GIRALT A, et al. Thermogenic Activation Induces FGF21 Expression and Release in Brown Adipose Tissue. J Biol Chem.2011;286(15):12983-12990.
[6] WU J, BOSTRÖM P, SPARKS L, et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell.2012; 150(2): 366-376.
[7] THYAGARAJAN B, FOSTER MT. Beiging of white adipose tissue as a therapeutic strategy for weight loss in humans. Horm Mol Biol Clin Investig.2017;31(2):0016.
[8] 张洁,牛燕媚,傅力.白色脂肪棕色化——运动减肥的新靶点[J].中国运动医学杂志, 2014,33(8):826-829+840.
[9] BARTELT A, HEEREN J. Adipose tissue browning and metabolic health. Nat Rev Endocrinol.2014;10(1):24-36.
[10] 高芸,庞卫军,杨公社.棕色和米色脂肪激活剂:潜在的减肥靶标[J].中国生物化学与分子生物学报, 2018, 34(4): 371-376.
[11] SEALE P, KAJIMURA S, YANG W, et al. Transcriptional Control of Brown Fat Determination by PRDM16. Cell Metab. 2007;6(1):0-54.
[12] TRAJKOVSKI M, AHMED K, ESAU CC, et al. MyomiR-133 regulates brown fat differentiation through Prdm16. Nat Cell Biol.2012;14(12): 1330-1335.
[13] SEO YJ, LEE K, CHEI S, et al. Ishige okamurae Extract Ameliorates the Hyperglycemia and Body Weight Gain of db/db Mice through Regulation of the PI3K/Akt Pathway and Thermogenic Factors by FGF21.Mar Drugs.2019;17(7).
[14] IZUMIYA Y, BINA HA, OUCHI N, et al. FGF21 is an Akt-regulated myokine. FEBS Letters. 2008;582(27):3805-3810.
[15] YAN Z, KRONEMBERGER A, BLOMME J, et al. Exercise leads to unfavourable cardiac remodelling and enhanced metabolic homeostasis in obese mice with cardiac and skeletal muscle autophagy deficiency. Sci Rep.2017;7:7894.
[16] PLANAVILA A, REDONDO I, HONDARES E, et al. Fibroblast growth factor 21 protects against cardiac hypertrophy in mice.Nat Commun. 2013;4(3):2019.
[17] YU J, YU B, JIANG H, et al. Conjugated linoleic acid induces hepatic expression of fibroblast growth factor 21 through PPARalpha.Br J Nutr.2012;107(4):461-465.
[18] YANG W, CHEN X, LIU Y, et al. N-3 polyunsaturated fatty acids increase hepatic fibroblast growth factor 21 sensitivity via aPPAR-gamma-beta-klotho pathway.Mol Nutr Food Res.2017; 61(9):1601075.
[19] LI H, WU G, FANG Q, et al. Fibroblast growth factor 21 increases insulin sensitivitythrough specific expansion of subcutaneous fat.Nat Commun.2018; 9(1):272.
[20] DESAI BN, SINGHAL G, WATANABE M, et al. Fibroblast growth factor 21 (FGF21) isrobustly induced by ethanol and has a protective role in ethanol associated liver injury. Mol Metab.2017;6(11):1395-1406.
[21] LIU Y, ZHAO C, XIAO J, et al. Fibroblast growth factor 21 deficiency exacerbates chronic alcohol-induced hepatic steatosis and injury. Sci Rep.2016;6:31026.
[22] BOOKOUT AL, DE GROOT MH, OWEN BM, et al. FGF21 regulates circadian behavior and metabolism by acting on the nervous system. Nat Med.2013;19(9):1147-1152.
[23] CHENG X, VISPUTE SG, LIU J, et al. Fibroblast growth factor (Fgf) 21 is a novel target gene of the aryl hydrocarbon receptor (AhR). Toxicol Appl Pharm. 2014;278(1):65-71.
[24] KEIPERT S, OST M, JOHANN K, et al. Skeletal muscle mitochondrial uncoupling drives endocrine cross-talk through the induction of FGF21 as a myokine. Am J Physiol Endocrinol Metab. 2014; 306(5):E469-482.
[25] VÉNIANT MURIELLE M, CLARENCE H, JOAN H, et al. FGF21 Promotes Metabolic Homeostasis via White Adipose and Leptin in Mice. PLoS One.2012;7(7):e40164.
[26] LIN Z, TIAN H, LAM KS, et al. Adiponectin mediates the metabolic effects of FGF21 on glucose homeostasis and insulin sensitivity inmice. Cell Metab.2013;17:779-789.
[27] HOLLAND WL, ADAMS AC, BROZINICK JT, et al. An FGF21-adiponectin-ceramide axis controls energy expenditure and insulin action in mice. Cell Metab.2013;17:790-797.
[28] CAO L, CHOI E, LIU X, et al. White to brown fat phenotypic switch induced by genetic and environmental activation of a hypothalamic- adipocyte axis.Cell Metab.2001;14(3):324-338.
[29] LEE P, SMITH S, LINDERMAN J, et al. Temperature-acclimated brown adipose tissue modulates insulin sensitivity in humans.Diabetes.2014; 63:3686-3698.
[30] FISHER FM, ESTALL JL, ADAMS AC, et al. Integratedregulation of hepatic metabolism by fibroblast growth factor 21 (FGF21) in vivo. Endocrinology. 2011;152(8):2996–3004.
[31] LEE P, LINDERMAN JD, SMITH S, et al. Irisin and FGF21 Are Cold-Induced Endocrine Activators of Brown Fat Function in Humans. Cell Metab.2014;19(2):302-309.
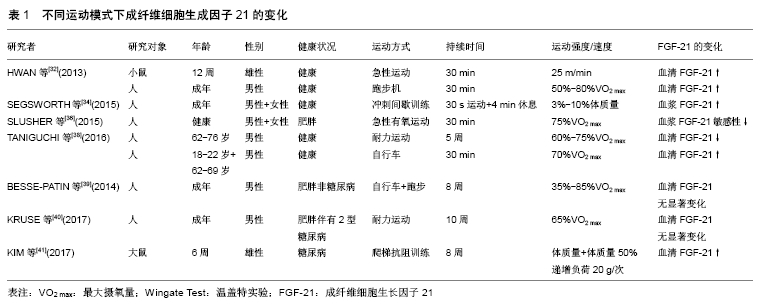
[32] HWAN KK, HUN KS, YOUNG-KI M, et al. Acute Exercise Induces FGF21 Expression in Mice and in Healthy Humans. PLoS One.2013; 8(5):e63517.
[33] DA SILVEIRA CAMPOS RM, DMASO AR,MASQUIO DCL, et al. The effects of exercise training associated with low-level laser therapy on biomarkers of adipose tissue transdifferentiation in obese women. Lasers Med Sci.2018;33:1245-1254.
[34] SEGSWORTH, BLAIR M. Acute Sprint Interval Exercise Induces a Greater FGF-21 Response in Comparison to Work-Matched Continuous Exercise. Elec Thesis Dis Rep.2015; 3254.
[35] FLETCHER JA, MEERS GM, LAUGHLIN MH, et al. Modulating fibroblast growth factor 21 in hyperphagic OLETF rats with daily exercise and caloric restriction. Appl Physiol Nutr Metab.2012; 337(6): 1054-1062.
[36] SLUSHER AL, WHITEHURST M, ZOELLER RF, et al. Attenuated fibroblast growth factor 21 response to acute aerobic exercise in obese individuals.Nutr Metab Cardiovasc Dis.2015; 25(9):839-845.
[37] TANIGUCHI H,TANISAWA K, SUN X, et al. Endurance exercise reduces hepatic fat content and serum fibroblast growth factor 21 levels in elderly men.J Clin Endocrinol Metab.2016;101(1):191.
[38] TANIGUCHI H, TANISAWA K, SUN X, et al. Acute endurance exercise lowers serum fibroblast growth factor 21 levels in Japanese men. Clin Endocrinol.2016;85(6): 861-867.
[39] BESSE-PATIN A, MONTASTIER E, VINEL C, et al. Effect of endurance training on skeletal muscle myokine expression in obese men: identification of apelin as a novel myokine.Int J Obes.2014;38(5): 707-713.
[40] KRUSE R, VIENBERG SG, VIND BF, et al. Effects of insulin and exercise training on FGF21, its receptors and target genes in obesity and type 2 diabetes. Diabetologia.2017;60(10):2042.
[41] KIM HJ, SONG W. Resistance training increases fibroblast growth factor-21 and irisin levels in the skeletal muscle of Zucker diabetic fatty rats. J Exerc Nutrition Biochem.2017;21(3):50-54.
[42] LEE P, LINDERMAN JD, SMITH S, et al. Irisin and FGF21 are cold-induced endocrine activators of brown fat function in humans. Cell Metab.2014;19: 302-309.
[43] KNUDSEN JG, MURHOLM M, CAREY AL, et al. Role of IL-6 in Exercise Training- and Cold-Induced UCP1 Expression in Subcutaneous White Adipose Tissue. PLoS One.2014;9(1):e84910.
[44] LI G, XIE C, LU S, et al. Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota.Cell Metab.2017;26(5):801.
[45] THYAGARAJAN B, FOSTER MT. Beiging of white adipose tissue as a therapeutic strategy for weight loss in humans. Horm Mol Biol Clin Investig.2017;31(2):1-13.
|