| [1] Vale RD. The molecular motor toolbox for intracellar transport. Cell. 2003;112: 467-470.
[2] Zhang HW,Wen ST, Zhang H , et al. A feedback- controlled Brownian ratchet operated by a temperature switch. Chin Phys B. 2012;21(7):078701.
[3] Hirokawa N. Organelle transport along microtubules- the role of KIFs. Trends Cell Biol. 1996;6(4):135-141.
[4] Hirokawa N, Noda Y, Okoda Y. Kinesin and dynein superfaimly proteins in organelle transport and cell division. Curr Opin Cell Biol. 1998;10(1):60-73.
[5] Hirokawa N, Takemura R. Molecular motors and mechanisms of directional transport in neurons. Nat Rev Neurosci. 2005;6: 201-214.
[6] Sharp DJ, Rogers GC, Seholey JM. Micotubule motors in mitosis. Nature.2000;407:41-47.
[7] Miki H, Okada Y, Hirokawa N. Analysis of kinesin superfaimly: insights into structure and function. Trends Cell Biol. 2005;15(9):467-476.
[8] Liu M, Xu N, Ruan S, et al. Resrarch Progress of Kinesin and Its Function. J Hangzhou Normal Univ. 2013;(1):40-44.
[9] Hirokawa N, Takemura R. Kinesin superfaimly proteins and their various functions and dynamics. Exp Cell Res. 2004;301, 50-59.
[10] Shi SH, Cheng T, Jan LY, et al. APC and GSK-3beta are involved in mPar3 targeting to the nascent axon and establishment of neuronal polarity. Curr Biol. 2004; 14: 2025-2032.
[11] Yamazaki H, Nakata T, Okada Y, et al. Cloning and characterization of KAP3: a novel kinesin superfamily- associated protein of KIF3A/3B. Proc Natl Acad Sci USA. 1996;93: 8443-8448.
[12] Whitehead JL, Wang SY, Bost-Usinger L, et al. Photoreceptor Localization of the KIF3A and KIF3B Subunits of the Heterotrimeric Microtubule Motor Kinesin II in Vertebrate Retina. Exp Eye Res. 1999;69: 491-503.
[13] Aizawa H, Sekine Y, Takemura R, et al. Kinesin family in murine central nervous system. Cell Biol. 1992;119: 1287-1296.
[14] Frazer KA, Ueda Y, Zhu Y, et al. Computational and biological analysis of 680 kb of DNA sequence from the human 5q31 cytokine gene cluster region. Gen Res. 1997;7:495-512.
[15] Makalowski W, Zhang J, Boguski MS. Comparative analysis of 1196 orthologous mouse and human full-length mRNA and protein sequences. Gen Res. 1996;(9):846-857.
[16] Goetz SC, Anderson KV. The primary cilium: a signalling centre during vertebrate development. Nat Rev Genet 2010;11: 331-344.
[17] Scholey JM, Anderson KV. Intraflagellar transport and cilium-based signaling. Cell. 2006;125: 439-442.
[18] Marszalek JR, Ruiz-Lozano P, Roberts E et al. Situs inversus and embryonic ciliary morphogenesis defects in mouse mutants lacking the KIF3A subunit of kinesin-II. Proc Natl Acad Sci USA. 1999;96: 5043-5048.
[19] Berbari NF, O’Connor AK, Haycraft CJ, et al. The primary cilium as a complex signaling center. Curr Biol. 2009; (19):526-535.
[20] Davis, EE, Brueckner M, Katsanis N. The emerging complexity of the vertebrate cilium: new functional roles for an ancient organelle. Dev Cell. 2006;(11): 9-19.
[21] Xiao Z, Zhang S, Mahlios J, et al. Cilia-like structures and polycystin-1 in osteoblasts/osteocytes and associated abnormalities in skeletogenesis and Runx2 expression. Biol Chem. 2006;(281): 30884-30895.
[22] Gerdes JM, Davis EE, Katsanis N. The vertebrate primary cilium in development, homeostasis, and disease. Cell. 2009;(137):32-45.
[23] Ni Q, Zhou SX, Li C, et al. Disruption of Kif3a in osteoblasts results in defective bone formation and osteopenia. J Cell Sci. 2012;(125):1945-1957.
[24] Christensen ST, Pedersen LB, Schneider L, et al. Sensory cilia and integration of signal transduction in human health and disease. Traffic. 2007;(8): 97-109.
[25] Han YG, Spassky N, Romaguera-Ros M, et al. Hedgehog signaling and primary cilia are required for the formation of adult neural stem cells. Nat Neurosci. 2008;(11):277-284.
[26] Huang FD, Anderson KV. Cilia and Hedgehog responsiveness in the mouse. Proc Natl Acad Sci USA. 2005;(102): 11325-11330.
[27] Kovacs JJ, Whalen EJ, Liu R, et al. Beta-arrestin- mediated localization of smoothened to the primary cilium.Science. 2008;( 320): 1777-1781.
[28] Serra R. Role of intraflagellar transport and primary cilia in skelet al development. Anat Rec (Hoboken). 2008;(291): 1049-1061.
[29] Veland IR, Awan A, Pedersen LB, et al. Primary cilia and signaling pathways in mammalian development, health and disease. Nephron Physiol. 2009;(111): 39-53.
[30] Wong SY, Seol AD, So PL, et al. Primary cilia can both mediate and suppress Hedgehog pathway-dependent tumorigenesis. Nat Med. 2009;(15): 1055-1061.
[31] Li Q, Montalbetti N, Wu Y, et al. Polycystin-2 cation channel function is under the control of microtubular structures in primary cilia of renal epithelial cells. Biol Chem. 2006;281:37566-37575.
[32] Xiao Z, Dallas M, Qiu N, et al. Conditional deletion of Pkd1 in osteocytes disrupts skeletal mechanosensing in mice. FASEB J. 2011;25: 2418-2432.
[33] Xiao Z, Zhang S, Cao L, et al. Conditional disruption of Pkd1 in osteoblasts results in osteopenia due to direct impairment of bone formation. Biol. Chem. 2010;285: 1177-1187.
[34] Haycraft CJ, Zhang Q, Song B, et al. Intraflagellar transport is essential for endochondral bone formation. Development. 2007;134:307-316.
[35] Kolpakova HE, Jinnin M, Hou B, et al. Kinesin-2 controls development and patterning of the vertebrate skeleton by Hedgehog- and Gli3-dependent mechanisms. Dev Biol. 2007;309: 273-284.
[36] Christopher B, Fruzsina K, Bjoern B, et al. Kif3a Guides Microtubular Dynamics, Migration and Lumen Formation of MDCK Cells. PLoS ONE. 2013;8(5): e62165.
[37] Huangfu D, Liu A, Rakeman AS, et al. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature. 2003;426: 83-87.
[38] Goetz SC, Anderson KV. The primary cilium: a signalling centre during vertebrate development. Nat Rev Genet. 2010;11: 331-344.
[39] Teng J, Rai T, Tanaka Y, et al. The KIF3 motor transports N-cadherin and organizes the developing neuroepithelium. Nat Cell Biol. 2005;7: 474-482.
[40] Jaulin F, Kreitzer G. KIF17 stabilizes microtubules and contributes to epithelial morphogenesis by acting at MT plus ends with EB1 and APC. J Cell Biol. 2010;190: 443-460.
[41] Song B, Haycraft CJ, Seo HS, et al. Development of the post-natal growth plate requires intraflagellar transport proteins. Dev Biol. 2007;305:202-216.
[42] Murcia NS, Richards WG, Yoder BK, et al. The Oak Ridge Polycystic Kidney (orpk) disease gene is required for left-right axis determination. Development. 2000;127: 2347-2355.
[43] Mari SL, Noora K, Anu S. KIF1-binding protein interacts with KIF3A in haploid male germ cells. Reproduction. 2015;(150): 209-216.
[44] Lehti MS, Kotaja N, Sironen A, et al. KIF3A is essential for sperm tail formation and manchette function. Mol Cell Endocrinol. 2013; 377:44-55.
[45] Scholey JM. Intraflagellar transport motors in cilia: moving along the cell’s antenna. J Cell Biol. 2008;180 23-29.
[46] Hu J, Xu N, Tan FQ, et al. Molecular characterization of a Kif3A-like kineain gene in the testis of the Chinese fire-bellied newt Cynops orientalis. Mol Biol Rep. 2012; 39(4):4207-4214.
[47] Wang YT, Mao H, Hou CC, et al. Characterization and expression pattern of Kifc1-like kinesin gene in the testis of the Macrobrachium nipponense with discussion of its relationship with atructure lamellar complex(LCx) and acroframosome(AFS). Mol Biol Rep. 2012;9(7):7591-7598.
[48] Zun L, Ryan ER, Zemin W. KIF3a Promotes Proliferation and Invasion via Wnt Signaling in Advanced Prostate Cancer. Mol Cancer Res. 2014;12(4); 491-503.
[49] Lan B, Hoang M, Loic PD, et al. Disruption of KIF3A in patient-derived glioblastoma cells: effects on ciliogenesis, hedgehog sensitivity, and tumorigenesis. Oncotarget. 2016;(7):7029-7043.
[50] Monique TB, Eric WH, Matthew PS. Kif3a is Necessary for Initiation and Maintenance of Medulloblastoma. Carcinogenesis. 2013;34(6):1382-1392.
[51] Li JC, Alevtina G, Lin C,et al. Kif3a Controls Murine Nephron Number Via GLI3 Repressor, Cell Survival, and Gene Expression in a Lineage-Specific Manner. PLOS ONE. 2013; 8: e65448.
[52] Yoder BK. Role of primary cilia in the pathogenesis of polycystic kidney disease. Am Soc Nephrol. 2007;18: 1381-1388.
[53] Lin F, Hiesberger T, Cordes K, et al. Kidneyspecific inactivation of the KIF3A subunit of kinesin-II inhibits renal ciliogenesis and produces polycystic kidney disease. Proc Natl Acad Sci USA. 2003; 100: 5286-5291.
[54] Simons M, Gloy J, Ganner A, et al. Inversin, the gene product mutated in nephronophthisis type II, functions as a molecular switch between Wnt signaling pathways. Nat Genet. 2005; 37: 537-543.
[55] Gerdes JM, Davis EE, Katsanis N. The vertebrate primary cilium in evelopment, homeostasis, and disease. Cell. 2009;137: 32-45.
[56] Jonassen JA, San Agustin J, Follit JA, et al. Deletion of IFT20 in the mouse kidney causes misorientation of the mitotic spindle and cystic kidney disease. J Cell Biol. 2008;183: 377-384.
[57] Ille F, Sommer L. Wnt signaling: multiple functions in neur development. Cell Mol Life Sci. 2005;62(10): 1100-1108.
[58] Corbit KC, Shyer AE, Dowdle WE, et al. Kif3a constrains β-catenin-dependent Wnt signalling through dual ciliary and non-ciliary mechanisms. Nat Cell Biol. 2008;10(1):70-76.
[59] Val RD, Milligan RA. The way things move: looking under the hood of molecular motor protein.. Science. 2002; 288(5463):88-95.
[60] Schliwa M, Woehlke G. Molecular motors. Nature. 2003;422(6933):759-765.
[61] Ocbina PJ, Aanderson KV. Intraflagellar transport cilia and mammalian Hedgehog signalling : analysis in mouse embryonic fibroblasts. Dev Dyn. 2008;37(8):2030-2038.
[62] Liu Z, Rebowe RE, Wang Z, et al. KIF3a promotes proliferation and invasion via Wnt signaling in advanced prostate cancer. Mol Cancer Res. 2014; 12(4):491-503. |
.jpg) 文题释义:
KIF3A:是异三聚体驱动蛋白2的亚基,属于N-驱动蛋白,微管正端导向的马达蛋白。异三聚体驱动蛋白2则由KIF3A、KIF3B两个亚基和运载蛋白KAP3 组成。KIF3A作为驱动蛋白2家族的一个重要成员,除了拥有驱动蛋白的一些共性(包括调节细胞内运输,介导微管运动等)之外,它也具有自己特异性的功能。
分子马达:是一类具有马达功能的蛋白质大分子,主要为细胞内物质的运输提供动力。细胞内某些细胞器、囊泡以及蛋白质等物质,在远距离运输的过程中需要依赖于微管运输的马达蛋白的协同作用。驱动蛋白则是基于微管的分子马达蛋白,主要参与细胞器、囊泡、mRNA以及蛋白质等的转运。驱动蛋白通过控制微管的动力学变化,进而调节细胞的动力学变化。
文题释义:
KIF3A:是异三聚体驱动蛋白2的亚基,属于N-驱动蛋白,微管正端导向的马达蛋白。异三聚体驱动蛋白2则由KIF3A、KIF3B两个亚基和运载蛋白KAP3 组成。KIF3A作为驱动蛋白2家族的一个重要成员,除了拥有驱动蛋白的一些共性(包括调节细胞内运输,介导微管运动等)之外,它也具有自己特异性的功能。
分子马达:是一类具有马达功能的蛋白质大分子,主要为细胞内物质的运输提供动力。细胞内某些细胞器、囊泡以及蛋白质等物质,在远距离运输的过程中需要依赖于微管运输的马达蛋白的协同作用。驱动蛋白则是基于微管的分子马达蛋白,主要参与细胞器、囊泡、mRNA以及蛋白质等的转运。驱动蛋白通过控制微管的动力学变化,进而调节细胞的动力学变化。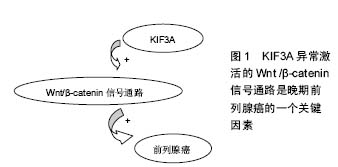
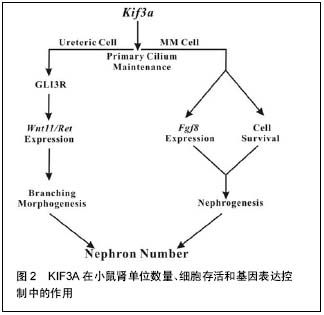
.jpg) 文题释义:
KIF3A:是异三聚体驱动蛋白2的亚基,属于N-驱动蛋白,微管正端导向的马达蛋白。异三聚体驱动蛋白2则由KIF3A、KIF3B两个亚基和运载蛋白KAP3 组成。KIF3A作为驱动蛋白2家族的一个重要成员,除了拥有驱动蛋白的一些共性(包括调节细胞内运输,介导微管运动等)之外,它也具有自己特异性的功能。
分子马达:是一类具有马达功能的蛋白质大分子,主要为细胞内物质的运输提供动力。细胞内某些细胞器、囊泡以及蛋白质等物质,在远距离运输的过程中需要依赖于微管运输的马达蛋白的协同作用。驱动蛋白则是基于微管的分子马达蛋白,主要参与细胞器、囊泡、mRNA以及蛋白质等的转运。驱动蛋白通过控制微管的动力学变化,进而调节细胞的动力学变化。
文题释义:
KIF3A:是异三聚体驱动蛋白2的亚基,属于N-驱动蛋白,微管正端导向的马达蛋白。异三聚体驱动蛋白2则由KIF3A、KIF3B两个亚基和运载蛋白KAP3 组成。KIF3A作为驱动蛋白2家族的一个重要成员,除了拥有驱动蛋白的一些共性(包括调节细胞内运输,介导微管运动等)之外,它也具有自己特异性的功能。
分子马达:是一类具有马达功能的蛋白质大分子,主要为细胞内物质的运输提供动力。细胞内某些细胞器、囊泡以及蛋白质等物质,在远距离运输的过程中需要依赖于微管运输的马达蛋白的协同作用。驱动蛋白则是基于微管的分子马达蛋白,主要参与细胞器、囊泡、mRNA以及蛋白质等的转运。驱动蛋白通过控制微管的动力学变化,进而调节细胞的动力学变化。