2.1 低氧环境下低氧诱导因子对骨骼的影响 低氧诱导因子是迄今为止发现的组织细胞在低氧状态下诱生的最直接或唯一的调控因子,包括3种亚型低氧诱导因子1、低氧诱导因子2和低氧诱导因子3[1]。低氧诱导因子1主要在骨髓和肝脏内表达,低氧诱导因子2主要在肺、心及肾脏内表达,低氧诱导因子3的调控作用主要集中在肺发育、脂肪细胞分化、耐力及脑膜瘤等中[2-5]。研究表明,低氧诱导因子2在成骨细胞分化中发挥负调控作用,抑制低氧诱导因子2将有利于低氧诱导因子1的表达水平[6-7]。目前发现低氧诱导因子对骨骼影响主要以低氧诱导因子1为主。
2.1.1 低氧诱导因子1通过血管内皮生长因子作用于骨骼 低氧诱导因子1是有低氧诱导因子1α和低氧诱导因子1β组成的异源二聚体[8],低氧环境可以促进低氧诱导因子1的生成。
在骨发育过程中,低氧诱导因子1α具有明显的促进血管生成及骨骼发育的作用。组织局部的缺氧是激活低氧诱导因子1α的一个重要诱因,低氧诱导因子1α可以调控血管内皮生长因子,血管内皮生长因子起到募集血管并使之侵入软骨的作用。Gerber等[9]研究发现,一旦激活低氧诱导因子1α通路,可以造成血管内皮生长因子和其他血管生成因子的过量合成、分泌,从而刺激长骨的血管生成。其主要过程为缺氧时低氧诱导因子1α先聚集,促进血管内皮生长因子的转录和表达,从而刺激大量相关血管生成因子的合成与分泌,而且血管内皮生长因子作用于血管内皮细胞表面的血管内皮生长因子受体,从而激活一系列的缺血转录通路,诱导新生血管生成。在软骨内成骨的过程中,肥大软骨细胞表达血管内皮生长因子的增加以及软骨膜周围血管内皮生长因子受体表达的增加导致血管侵入到软骨中,从而完成软骨化骨过程。当低氧诱导因子1α基因被移除后,小鼠在软骨内成骨过程中血管内皮生长因子表达减少,新骨形成速度减慢,钙元素含量和骨小梁面积减少[10]。
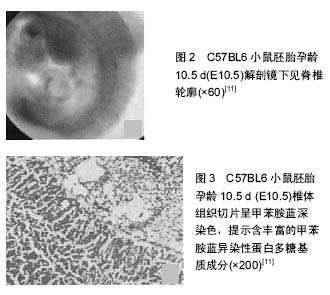
朱勋兵等[11]研究发现,在低氧环境下小鼠椎体发育伴有低氧诱导因子1αmRNA及蛋白量的增加及其下游基因血管内皮生长因子表达的增加。这一结果提示,低氧环境能够促进骨组织内血管生成,促进骨骼生长(见图2,3)。
血管内皮生长因子的细胞合成不仅受到低氧因素的诱导还受到其他信号通路的影响,如前列腺素 E1/E2、转化生长因子β、骨形态发生蛋白、成纤维细胞生长因子2、类胰岛素生长因子1、内皮素1和维生素D3,而这些蛋白因子也均受到低氧诱导因子1α通路的调控或影响。
2.1.2 低氧诱导因子1直接作用于骨骼 一方面低氧诱导因子1α可以通过血管内皮生长因子而促进骨骼生长,另一方面也可以直接作用于成骨细胞和破骨细胞。但是对于低氧诱导因子1α对于成骨细胞和破骨细胞的作用,研究结果却不一而论。
低氧坏境对成骨细胞的作用:低氧抑制成骨细胞的生成与分化。持续12 h低氧使低氧诱导因子1α中的下游基因Twist表达上调,抑制了Runx2下游靶基因的表达。Runx2是成骨细胞生成的一个主控基因,阻碍了骨髓间充质干细胞向成骨的分化,不利于成骨细胞的生成,此外,当氧浓度过低时(<1% O2)也不利于成骨细胞的生成[12]。成骨细胞是由具有多向分化潜能的间充质干细胞在特定转录因子诱导下分化而来的,低氧直接抑制了间充质干细胞的生成[13]。成骨细胞在低氧条件下(1% O2)培养36 h后,成骨细胞增殖细胞核抗原(PCNA,osteoblast pcna)的表达明显下降。低氧使低氧诱导因子1α稳定表达,明显抑制成骨细胞线粒体活性细胞增殖和骨钙素及成骨细胞表达骨保护素(osteoprotegerin,OPG)的表达。低氧的影响主要发生在成骨细胞分化的早期阶段,此阶段低氧可使成骨细胞处于过早分化状态,无法为基质的成熟和矿化提供正确信号[14]。
低氧能够促进成骨细胞的生成与分化。血管生成和骨形成在骨骼发育和骨折愈合中相互关联,血管内皮生长因子的表达能够促进成骨细胞的生成。成骨细胞在低氧处理3 h后,转化生长因子β1表达成倍增加,低氧9 h后血管内皮生长因子表达到达最高峰,随低氧时间延长,转化生长因子β1和血管内皮生长因子表达下降。短期低氧可能促进成骨细胞早期增殖及分化,长期低氧则可能抑制成骨细胞的增殖及分化[15]。此外,随着氧分压的降低,可以增强Runx2、骨形态发生蛋白2 mRNA和蛋白的表达,促进成骨细胞的生成[16]。也学者认为,低氧时低氧诱导因子1激活经典的Wnt/β-catenin通路,从而促进成骨细胞生长和分化[17]。Tseng等[18]研究发现,短期低氧能够通过低氧诱导因子1α,整合素连接激酶(ILK)、蛋白激酶B(Akt)和哺乳动物雷帕霉素靶蛋白(mTOR)途径诱导成骨细胞骨形态发生蛋白2的表达,促进成骨细胞分化。
低氧环境对破骨细胞的作用:目前有关低氧诱导因子1对破骨细胞的直接作用的研究还比较少,且未见明显作用。Wang等[19]研究发现,低氧诱导因子1α与低氧诱导因子2α的高表达并没有影响破骨细胞的数量与活性。在低氧条件下,诱导RANKL/骨保护素通路的比例发生变化时,骨保护素的表达量升高,骨吸收的活性在下降[20],破骨细胞的数量与活性也明显被抑制。此外,Yao等[21]研究发现,血管内皮生长因子也不能单独促进破骨细胞的形成与分化。
低氧环境下低氧诱导因子1主要是通过酸化作用刺激破骨细胞,而增加骨吸收能力。研究发现,低氧增加破骨细胞的生成与破骨细胞的分化阶段无关,而只是取决于低氧的持续时间。低氧时无氧代谢增加,酸性代谢产物积聚,可引起局部微环境轻度酸化,而破骨细胞的激活正是依赖于细胞外酸化[22]。常氧下(20%O2)培养破骨细胞形成,逐渐降低时,破骨细胞的数量、体积和骨吸收能力增加,最大的效应发生在2%O2时[23]。
有关低氧诱导因子对骨骼影响机制十分复杂,研究结果也不尽相同。不同的氧浓度下,低氧诱导因子对成骨细胞、破骨细胞的作用不一;在低氧环境下,成骨细胞及破骨细胞受到其他多种因素,如酸碱度、渗透压等;低氧诱导因子通过不同的通路及因子对骨骼的影响也不同。
2.2 低氧环境对骨代谢相关通路及其靶点的影响 在低氧环境下,骨代谢相关通路也有所改变。MAPK信号转导途径在成骨细胞分化过程中发挥着重要的作用,研究发现,在MAPK家族中,ERK、p38和JNK都参与了成骨细胞的分化。ERK1/2信号通路诱导Cbfa-1和ALP的表达,也可使基质矿化增强、向成骨方向分化速率加快[24]。此外,Minet等[25]研究发现,MAPK信号转导途径可以提高低氧诱导因子1α的转录活性,其原因是低氧诱导因子1α/低氧诱导因子2α在低氧状态下的调控都需要MAPK p42/44或PI3K的存在。
Wnt信号通路又称Wnt/β-catenin,是骨发育过程中一条经典的通路,其对成骨细胞的作用广泛且复杂,既可以促进又可以抑制,主要取决于个体发育所处的时期[26]。当稳定表达Wnt1和Wnt3α时,可以诱导ALP的活性,促进成骨细胞的早期阶段分化,同时Wnt1和Wnt10可以抑制脂肪细胞分化,促进其向成骨细胞分化[27]。而Wnt5α介导的Wnt/Ca2+信号通路可以调节破骨细胞的分化与功能活性[28]。离体细胞实验中,发现低氧诱导增强Wnt信号通路,而缺氧条件下,则抑制Wnt信号通路。因此,推测不同的低氧浓度对于Wnt信号通路影响较大。
转化生长因子β/骨形态发生蛋白信号通路在细胞增殖、分化和细胞基质形成方面也发挥着比较广泛而重要的功能,并且MAPK和Wnt信号可以对其进行调控[29]。Kale等[30]发现,转化生长因子β1能引起细胞积聚,刺激成骨生成,并提高ALP、I型胶原及骨钙素的活性,而转化生长因子β3则会显著降低ALP的表达。造成这种结果,可能是与转化生长因子β/骨形态发生蛋白信号通路被激活的程度及时间有关。
RANKL/RANK/OPG信号通路在成骨细胞及破骨细胞的通讯中起着关键的作用,成骨细胞通过改变骨保护素和RANKL的合成,从而间接调节破骨细胞的分化和成熟。RANK结合RANKL能够促进破骨细胞的分化及成熟,但在这个过程中,骨保护素作为一种诱饵受体,可以阻断RANK和RANKL的结合,骨吸收和骨形成达到平衡的关键就在于RANKL/OPG的比值[31]。Yang等[20]研究发现,在低氧条件下,RANKL/RANK/OPG信号通路能够促进成骨细胞的生成,对破骨细胞则没有影响。原因可能是低氧环境诱导了骨保护素的水平增加,从而阻断了RANK和RANKL的结合,抑制了破骨细胞的分化和成熟。
Notch信号对骨的发育及骨细胞的活性很重要,调节成骨细胞和破骨细胞的分化和功能,它的作用与细胞环境相关,不同的氧浓度对Notch信号通路的影响不同[32-34]。当Notch信号通路被激活时,初始阶段主要由Notch1促进成骨细胞的分化及成熟,到了后期主要由Notch2提高了破骨细胞的活性。Hilton等[35]的研究表明,在骨髓间充质干细胞中当Notch信号通路被激活时,8周时会出现骨量增加,但在26周时变异组的骨量减少到对照组的10%。
不同的骨代谢相关信号通路在低氧环境下被激活的程度不一,成骨细胞介导的骨形成和破骨细胞介导的骨吸收平衡随之发生改变,随着时间的持续,对骨生长及重建的效果也不一。
2.3 低氧环境下运动训练对骨骼的影响 首先,运动训练对骨代谢的影响是显而易见的。研究显示,钙、激素和维生素D等在骨代谢方面只是具有3%-10%的作用。而更多的则是力学的应力,达到了40%以上[36]。适宜的运动强度对一些骨生化指标,如钙、镁、磷及碱性磷酸酶、甲状旁腺激素等有一定的影响,能够促进骨密度的增大、骨量的累积,促进骨的形成[37-41]。此外,不同的运动项目对骨代谢的影响也不尽相同。由于不同运动项目对于人体骨骼系统的应力作用不同,导致运动项目与骨密度和骨量累积的选择性有一定差异。一般认为,篮球、足球、网球等冲击力较大的运动项目要比行走等冲击力小的运动项目更有效地增加骨密度和累计骨量;陆地上的运动项目要比水中的运动项目更有利于骨密度的增加和骨量的累积[42-44]。
较低的运动强度对骨代谢的影响是基本上没有促进的作用,过大的强度或过度的运动则会对骨代谢造成不利的影响,可以导致骨量的流失,甚至疲劳性骨折等问题。尤其对于女性运动员,研究发现,过度的运动导致年轻女性运动员雌激素缺乏,致使骨量流失,在闭经期的女性运动员,如果过度运动,其骨折的发生率会明显增加[45]。
在低氧环境下,对骨生长及骨重建的影响本身就比较复杂,骨生长及重建在低氧环境中持续的时间和不同的氧浓度条件下,表型不一。此外,低氧环境下,是否会与运动训练对骨代谢影响的一些骨生化指标和机制产生矛盾。比如,在缺氧的情况下,机体的血钙水平会下降,且随着缺氧程度的增加而下降的更加明显。因此,在低氧环境下进行运动训练,可能会导致骨量流失,不利于骨生长发育。张红品等[46]在研究中,就提出高原训练模式对少儿骨骼发育不利的说法。
在低氧环境下,目前的研究主要都是通过低氧诱导因子、与成骨细胞和破骨细胞相关信号通路对骨骼产生影响。短期内,低氧诱导因子1α能够促进成骨细胞的分化,随着时间慢慢增长,这种作用将逐渐减少,长期低氧诱导因子1α则会抑制成骨细胞的分化。长期低氧环境中,低氧诱导因子1α对破骨细胞作用不明显,主要通过酸化作用能刺激破骨细胞,促进骨吸收。相关信号通路在低氧环境下被激活后也主要是与其作用时间及氧浓度有关,其作用也不一。因此,根据低氧环境中低氧诱导因子及相关信号通路对成骨细胞及破骨细胞影响中推测:低氧环境下训练在刚开始阶段能够促进骨的生长及骨量的累积,在持续低氧环境中,训练时间增长会不利于骨的生长及骨量的累积,到后期抑制骨的生长及骨量的累积,导致骨发育迟缓或者骨质疏松等骨疾病。当氧浓度慢慢下降,降至某个临界值的时候,对骨的生长及骨量累积也都将产生不利影响。此过程的运动训练对骨代谢的影响存在诸多不可知的因素,运动训练本身要求要有适宜的强度,并且不能过度训练。假设在低氧环境下,一旦出现强度过大或者过度训练,是否会对运动员的骨代谢造成双重的不利影响。对于女性运动员,在闭经期是否可以进行低氧环境下的运动训练。低氧环境下对骨代谢的影响和运动训练对骨代谢的影响,它们之间组合对骨代谢又将会产生何种影响。此外,低氧环境是否会对机体酸碱度、渗透压等水平产生影响,从而影响到骨生长及骨量的累积。
.jpg) 文题释义:
低氧:是指高原环境随海拔增高, 形成大气压下降和其中的氧分压降低, 即空气中的氧含量低下, 导致吸入气氧分压下降而引起人体的低氧血症。
骨代谢:骨的功能是为肌肉收缩提供附着处及保护内脏等重要的生命器官。一般认为骨在细胞水平上是不活跃的,事实上骨的细胞在不停地进行着细胞代谢,不仅骨的细胞之间会相互作用,还存在骨髓中的红细胞生成细胞、基质细胞相互作用,以进行骨的改建和重建。有两种细胞在骨代谢中起着重要的作用,一种是吸收骨基质的破骨细胞,另一种是合成骨基质的成骨细胞。该两者分布在骨膜、骨小梁及骨皮质处。在两种细胞相互作用的部位被称作为基本多细胞单位。在每一个基本多细胞单位,骨可因破骨细胞的吸收而消失,也能被重新合成骨的成骨细胞所取代。有些成骨细胞被掺合在骨基质中变成骨细胞。
文题释义:
低氧:是指高原环境随海拔增高, 形成大气压下降和其中的氧分压降低, 即空气中的氧含量低下, 导致吸入气氧分压下降而引起人体的低氧血症。
骨代谢:骨的功能是为肌肉收缩提供附着处及保护内脏等重要的生命器官。一般认为骨在细胞水平上是不活跃的,事实上骨的细胞在不停地进行着细胞代谢,不仅骨的细胞之间会相互作用,还存在骨髓中的红细胞生成细胞、基质细胞相互作用,以进行骨的改建和重建。有两种细胞在骨代谢中起着重要的作用,一种是吸收骨基质的破骨细胞,另一种是合成骨基质的成骨细胞。该两者分布在骨膜、骨小梁及骨皮质处。在两种细胞相互作用的部位被称作为基本多细胞单位。在每一个基本多细胞单位,骨可因破骨细胞的吸收而消失,也能被重新合成骨的成骨细胞所取代。有些成骨细胞被掺合在骨基质中变成骨细胞。.jpg)
.jpg) 文题释义:
低氧:是指高原环境随海拔增高, 形成大气压下降和其中的氧分压降低, 即空气中的氧含量低下, 导致吸入气氧分压下降而引起人体的低氧血症。
骨代谢:骨的功能是为肌肉收缩提供附着处及保护内脏等重要的生命器官。一般认为骨在细胞水平上是不活跃的,事实上骨的细胞在不停地进行着细胞代谢,不仅骨的细胞之间会相互作用,还存在骨髓中的红细胞生成细胞、基质细胞相互作用,以进行骨的改建和重建。有两种细胞在骨代谢中起着重要的作用,一种是吸收骨基质的破骨细胞,另一种是合成骨基质的成骨细胞。该两者分布在骨膜、骨小梁及骨皮质处。在两种细胞相互作用的部位被称作为基本多细胞单位。在每一个基本多细胞单位,骨可因破骨细胞的吸收而消失,也能被重新合成骨的成骨细胞所取代。有些成骨细胞被掺合在骨基质中变成骨细胞。
文题释义:
低氧:是指高原环境随海拔增高, 形成大气压下降和其中的氧分压降低, 即空气中的氧含量低下, 导致吸入气氧分压下降而引起人体的低氧血症。
骨代谢:骨的功能是为肌肉收缩提供附着处及保护内脏等重要的生命器官。一般认为骨在细胞水平上是不活跃的,事实上骨的细胞在不停地进行着细胞代谢,不仅骨的细胞之间会相互作用,还存在骨髓中的红细胞生成细胞、基质细胞相互作用,以进行骨的改建和重建。有两种细胞在骨代谢中起着重要的作用,一种是吸收骨基质的破骨细胞,另一种是合成骨基质的成骨细胞。该两者分布在骨膜、骨小梁及骨皮质处。在两种细胞相互作用的部位被称作为基本多细胞单位。在每一个基本多细胞单位,骨可因破骨细胞的吸收而消失,也能被重新合成骨的成骨细胞所取代。有些成骨细胞被掺合在骨基质中变成骨细胞。