3.1 纳入文献研究的结果分析 骨保护素、细胞核因子κB受体活化因子配体(RANKL)以及细胞核因子κB受体活化因子(RANK)组成的骨保护素-RANKL-RANK系统是近年来发现的在破骨细胞分化过程中的一个重要的信号传导通路,具有调控破骨细胞分化成熟的作用。RANKL与破骨细胞膜上特异性受体RANK结合,促进破骨细胞分化,增强成熟破骨细胞的活力,抑制破骨细胞凋亡。骨保护素通过阻断RANKL与RANK结合而抑制前体破骨细胞分化,抑制成熟破骨细胞骨吸收活性,促进破骨细胞凋亡[6]。
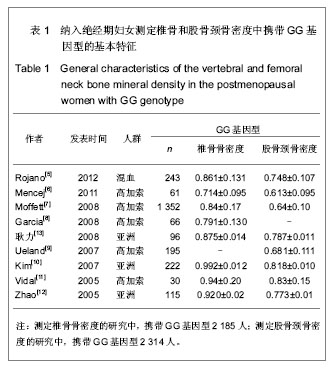
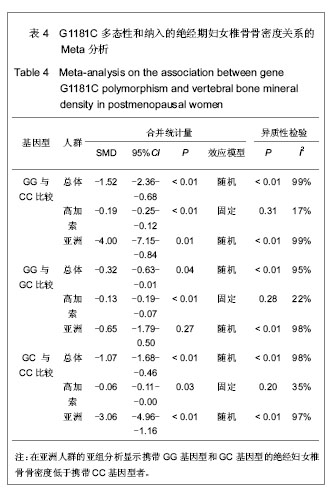
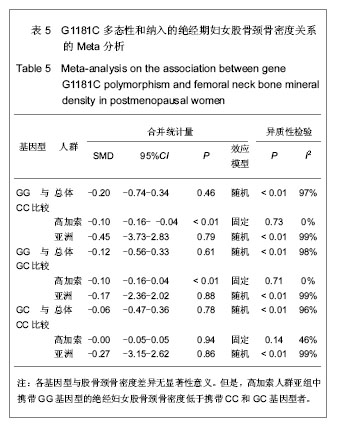
由于骨保护素在骨代谢中的重要作用,骨保护素基因被认为是骨质疏松症的候选基因[5]。G1181C多态性(rs2073618)于2002年对爱尔兰绝经前后妇女骨保护素基因编码区筛查中首次发现。此后大多数研究结果均提示G1181C与骨密度有关,包括对205例中国绝经妇女的研究发现,含G等位基因者椎骨骨密度更低,发生绝经后骨质疏松症的风险较C等位基因高2.7倍[16]。对美国 6 640例65岁及以上年龄妇女的研究同样发现,含G等位基因者骨密度低于含C等位基因者[10]。但是,对马耳他绝经妇女及澳大利亚老年妇女的研究却发现G1181C与骨密度无关[12-13]。Lee等[4]曾使用Meta分析方法研究过G1181C多态性与骨密度之间的关系,但是该研究尚存在不足之处,包括:①纳入的对象人群混杂,既包括绝经前和绝经后妇女,又包括男性骨质疏松患者,而未进行有效的分层分析。②由于Lee等的研究进行时间较早且受到语言限制,所纳入的针对亚洲人群,特别是中国人群的文献较少。所以,作者再次对G1181C多态性与骨密度之间的关系进行Meta分析。首先,作者缩小了对象人群范围,限定在绝经后妇女,这样可以最大程度上避免潜在混杂因素对结果的影响。其次,随着近年来该领域研究文献数量的增加,使得所纳入的针对亚洲人群,特别是中国人群的相关研究得到补充。最终,本实验对9篇符合纳入标准的文献进行了Meta分析。结果显示,在基因型与椎体骨密度关系中,携带G等位基因的绝经妇女椎体骨密度比携带C等位基因者低,高加索人群亚组分析显示相同结果,亚洲人群亚组分析显示携带GG基因型和GC基因型的绝经妇女的椎骨骨密度低于携带CC基因型者。在基因型与股骨颈骨密度关系中,虽然总体分析显示不同基因型间股骨颈骨密度差异没有统计学意义,但是高加索人群的亚组分析显示,携带GG基因型的绝经妇女股骨颈骨密度低于携带GC和CC基因型者。作者认为,亚洲人群的亚组分析结果未发现统计学意义的原因可能是由于目前相关研究尚少,与研究对象样本数量小有关。
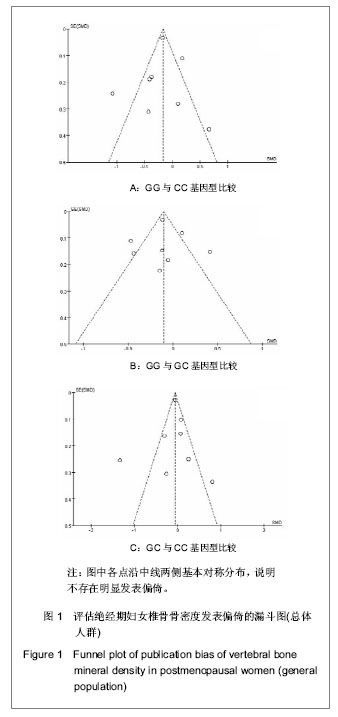
3.2 本研究的局限性 Meta分析结果的偏倚与原始研究的发表偏倚、选择偏倚和抽样偏倚有关。这次研究在资料收集阶段,在多个数据库中对相关文献进行检索,尽可能全面地检索所有相关文献,并采用漏斗图分析是否存在发表偏倚,但仍可能因部分阴性结果未能公开发表而具有潜在的发表偏倚。本研究在文献筛选阶段,制定了严格的文献纳入和排除标准,剔除了不合格文献;在数据提取阶段,采用两人独立提取,并进行核对,尽量避免数据提取的选择偏倚和人为错误。此外,对纳入Meta分析的各研究进行Hardy-Weinberg遗传平衡检验,发现有1篇文献的样本人群基因型分布不符合遗传平衡规律,表明该研究样本的随机性不够或样本量不足,所以纳入的研究仍可能存在抽样偏倚。但是,这次研究骨保护素基因G1181C多态性与绝经后妇女骨质密度关系Meta分析的结果还是为作者确定骨质疏松易感人群及提出综合防治措施提供了有力的依据。
3.3 结论与展望 综上所述,骨保护素 G1181C多态性确实与骨密度有关,其分子生物学机制是需要深入研究探讨的。骨保护素基因于1997年被发现,其编码产物属于肿瘤坏死因子受体(tumor necrosis factor receptor,TNFR)超家族成员,是一种分泌型糖蛋白。过度表达该蛋白的转基因小鼠骨质坚硬,呈明显的大理石样骨病表现;相反,敲除该基因的小鼠会发生严重的骨质疏松
[18]。骨保护素基因位于染色体8q23-24,含有5个外显子。G1181C多态性位于骨保护素基因第1外显子,第1外显子编码的是信号肽部分,是骨保护素从细胞中分泌所必需的。这一多态性使该外显子的第3个氨基酸由赖氨酸Lys(AAG)变成天冬氨酸Asn(AAC)
[7]。有学者认为,这种在基因信号肽区域的点突变可能会影响骨保护素的分泌动力学
[19- 20]。然而,针对不同地区人群的研究发现,G1181C多态性与血清骨保护素水平相关性无统计学意义
[9, 12, 15, 16]。所以,骨保护素基因G1181C多态性与骨密度关系的分子学机制还有待更多更深入的研究。