2.1 实验动物数量分析 共购入40只大鼠,实验分组所需36只,其中24只需建立急性脊髓损伤模型,造模过程中死亡3只,用多购入的4只补充,剩余1只未做处理,实验结束后处死。造模成功24只,造模成功率为88.9%,实验过程顺利,死亡3只大鼠死因均为造模时大量失血,所有大鼠均在灌胃后14 d后处死取材。
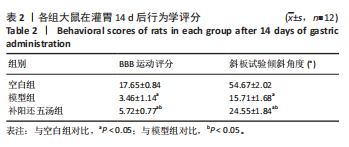
2.2 行为学评分结果 灌胃14 d后,各组大鼠的行为学评分见表2,与空白组对比,模型组大鼠下肢BBB运动评分明显较低
(P < 0.001),斜板试验倾斜角度明显较小(P < 0.001)。与模型组对比,补阳还五汤组大鼠下肢BBB运动评分较高(P < 0.05);斜板试验倾斜角度明显较大(P < 0.001)。
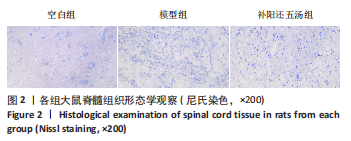
2.3 形态学观察结果 尼氏染色结果显示,空白组可见形态结构相对正常的神经元细胞结构完整,间隙正常,可见清晰的核仁核膜;模型组细胞坏死严重,结构紊乱,细胞核固缩,大部分核仁、核膜消失,存在大量组织空洞,有炎性细胞浸润;补阳还五汤组可见形态不规则、结构相对完整的神经元细胞,肿胀较轻,组织空洞和细胞坏死减少,见图2。
2.4 蛋白组学分析结果
2.4.1 差异蛋白的统计分析 研究共识别出54 412个肽段,其中40 229个为独特肽段,鉴定到7 563种蛋白质,其中7 546种蛋白质可进行定量分析。这些蛋白质在GO中注释了7 199条功能条目,参与了3 676条KEGG通路,并在蛋白质相邻类的聚簇(cluster of orthologous groups of proteins,COG)/真核型同源群(eukaryotic orthologous groups,KOG)功能分类中涵盖了6 305条功能类别。
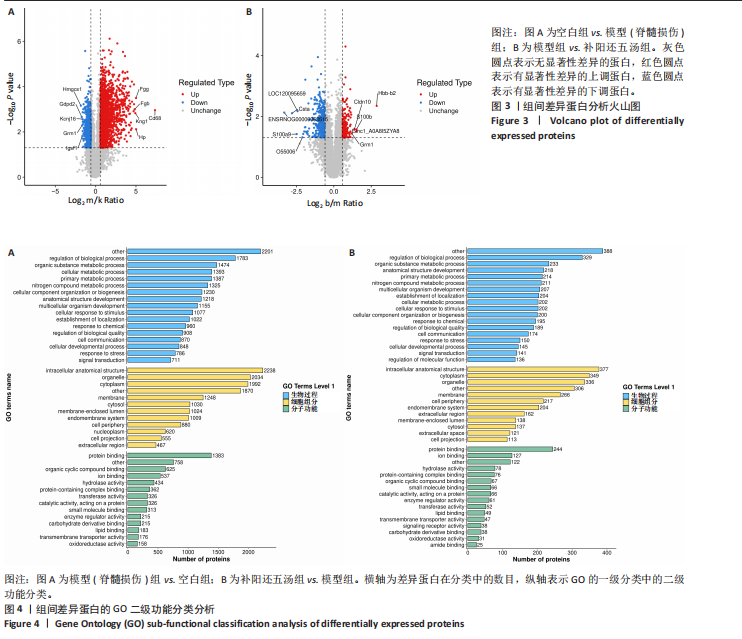
在模型组与空白组的比较中,共鉴定出2 541个差异表达蛋白质,其中1 774个蛋白质表达水平显著升高,767个蛋白质表达水平显著降低。选取特异性最强的前10位蛋白进行可视化分析,它们分别为Hmgcs1、Gdpd2、Kcnj16、Grm1、Igsf1、Fgg、Fgb、Cd68、Kng1、Hp,见图3A。在补阳还五汤组与模型组的对比中,共识别出496个差异表达蛋白,其中190个蛋白表达量显著上升,306个蛋白表达量显著下降。选取特异性最强的前10位蛋白进行可视化分析,它们分别为LOC120095659、Csta、ENSRNOG00000063615、S100a9、O55006、Hbb-b2、Cldn10、S100b、Emc1_A0A815ZYA8、Grm1,见图3B。
2.4.2 GO二级功能分类 GO二级功能分类包括生物过程(biological process,BP)、细胞组分(cellular component,CC)和分子功能(molecular function,MF)3个方面。
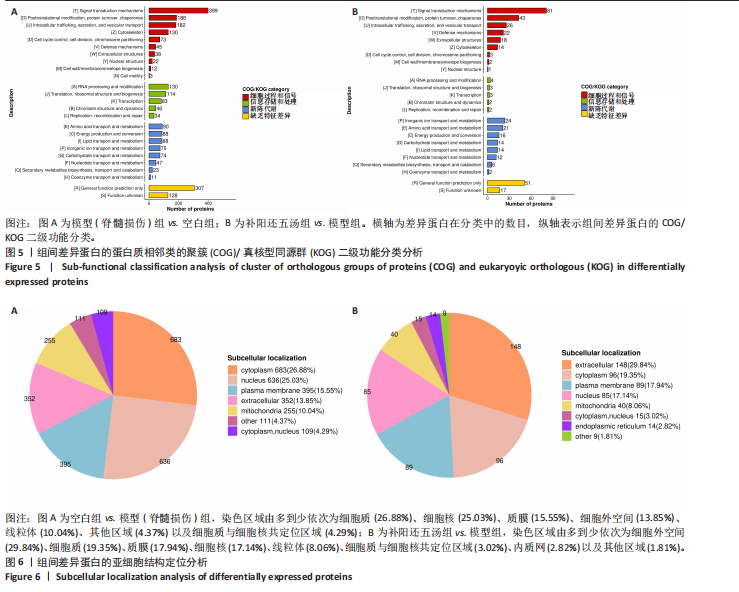
(1)生物过程分析:与空白组相比,模型组的差异表达蛋白主要参与生物过程的调控、有机物质代谢、细胞代谢、初级代谢、氮化合物代谢、细胞组分组织或生物发生、解剖结构发育、多细胞生物发育、细胞对刺激的反应、定位建立、对化学物质的反应、生物质量调控、细胞通信、细胞发育过程、对压力的反应以及信号传导等过程(见图4A)。与模型组相比,补阳还五汤组的差异蛋白则集中在生物过程的调控、有机物质代谢、细胞代谢、初级代谢、氮化合物代谢、细胞组分组织或生物发生、解剖结构发育、多细胞生物发育、细胞对刺激的反应、定位建立、对化学物质的反应、生物质量调控、细胞通信、细胞发育过程、对压力的反应、信号传导以及分子功能调控等生物过程(见图4B)。总体而言,两个对比组之间的差异蛋白均显著富集于上述15个生物学过程,但富集的显著性有所不同。
(2)细胞组分分析:与空白组相比,模型组的差异蛋白主要分布在细胞内解剖结构、细胞器、细胞质、膜、胞质、膜包裹的空腔、内膜系统、细胞周缘、核质、细胞投影以及胞外区域等细胞组分中(见图4A)。与模型组相比,补阳还五汤组的差异蛋白则主要集中在细胞内解剖结构、细胞器、细胞质、膜、胞质、膜包裹的空腔、内膜系统、胞外区域、细胞周缘、胞外间隙以及细胞投影等细胞组分中(见图4B)。总体而言,两个对比组之间的差异蛋白均显著富集于上述12个细胞组分,但富集的显著性有所不同。
(3)分子功能分析:与空白组相比,模型组的差异表达蛋白主要涉及细胞质结合、有机环状化合物结合、水解酶活性、离子结合、含蛋白质复合物结合、转移酶活性、蛋白催化活性、小分子结合、酶调节剂活性、碳水化合物衍生物结合、脂质结合、跨膜转运蛋白活性以及氧化还原酶活性等分子功能(见图4A)。与模型组相比,补阳还五汤组的差异蛋白也主要分布在细胞质结合、有机环状化合物结合、水解酶活性、离子结合、含蛋白质复合物结合、转移酶活性、蛋白催化活性、小分子结合、酶调节剂活性、碳水化合物衍生物结合、脂质结合、跨膜转运蛋白活性、氧化还原酶活性、信号受体活性以及酰胺结合等分子功能(见图4B)。总体而言,两个对比组之间的差异蛋白均显著富集于上述14个分子功能,但富集的显著性有所不同。
2.4.3 同源蛋白族分类分析 蛋白质相邻类的聚簇/真核型同源群分类涵盖了细胞过程和信号、信息存储和处理、新陈代谢以及缺乏特征差异等功能类别。
(1)细胞过程和信号分析:与空白组筛选到的差异蛋白相比,模型组的差异蛋白主要存在于信号转导机制、蛋白质转换和伴侣、细胞内运输、分泌和囊泡运输、细胞骨架、细胞周期控制、
细胞分裂和染色体分配、防御机制、细胞外结构、核结构、细胞壁/膜/囊泡生物发生以及细胞运动等方面(见图5A)。与模型组相比,补阳还五汤组的差异蛋白则主要集中在信号转导机制、蛋白质转换和伴侣、细胞内运输、细胞周期控制、分泌和囊泡运输、细胞骨架、细胞分裂和染色体分配、防御机制、细胞外结构、核结构以及细胞壁/膜/囊泡生物发生等细胞过程和信号通路中(见图5B)。总体而言,两个对比组之间的差异蛋白主要富集于上述9种细胞过程及信号通路,但富集程度存在差异。
(2)信息存储和处理分析:与空白组富集到的差异蛋白相比,模型组的差异蛋白主要涉及RNA加工和修饰、复制、重组和修复、翻译、核糖体结构和生物发生、染色质结构和动态以及转录等功能(见图5A)。与模型组相比,补阳还五汤组的差异蛋白则主要集中在复制、重组和修复、翻译、核糖体结构和生物发生、染色质结构和动态以及转录等信息存储和处理过程中(见图5B)。总体而言,两个对比组之间的差异蛋白主要富集于转录、翻译、核糖体结构和生物发生等4种信息存储和处理功能,但其富集程度有所不同。
(3)新陈代谢分析:与空白组相比,模型组富集到的差异蛋白主要参与氨基酸运输和代谢、能量生产和转换、脂质运输和代谢、无机离子运输和代谢、碳水化合物运输和代谢、核苷酸运输和代谢、次级代谢物的生物合成、运输和分解代谢以及辅酶运输和代谢等过程(见图5A)。与模型组相比,补阳还五汤组的差异蛋白则主要集中在氨基酸运输和代谢、能量生产和转换、无机离子运输和代谢、碳水化合物运输和代谢、核苷酸运输和代谢、次级代谢物的生物合成、运输和分解代谢以及辅酶运输和代谢等代谢过程中(见图5B)。总体而言,两个对比组之间的差异蛋白主要富集于上述7种新陈代谢过程,但每个过程过程富集程度不同。
(4)缺乏特征差异分析:两个对比组之间的差异蛋白均涉及未知功能类别。
2.4.4 亚细胞结构定位分析 与空白组相比,模型组亚细胞结构定位分析中的差异蛋白主要分布在细胞质(26.88%)、细胞核(25.03%)、质膜(15.55%)、细胞外空间(13.85%)、线粒体(10.04%)、其他区域(4.37%)以及细胞质与细胞核共定位区域(4.29%),如图6A所示。而与模型组相比,补阳还五汤组亚细胞结构定位分析中的差异蛋白则主要存在于细胞外空间(29.84%)、细胞质(19.35%)、质膜(17.94%)、细胞核(17.14%)、线粒体(8.06%)、细胞质与细胞核共定位区域(3.02%)、内质网(2.82%)以及其他区域(1.81%),如图6B所示。
总体而言,两个对比组之间的差异蛋白主要集中于细胞质、细胞外空间和线粒体等亚细胞结构,但其具体的定位分布及富集程度存在显著差异。
2.4.5 GO富集分析
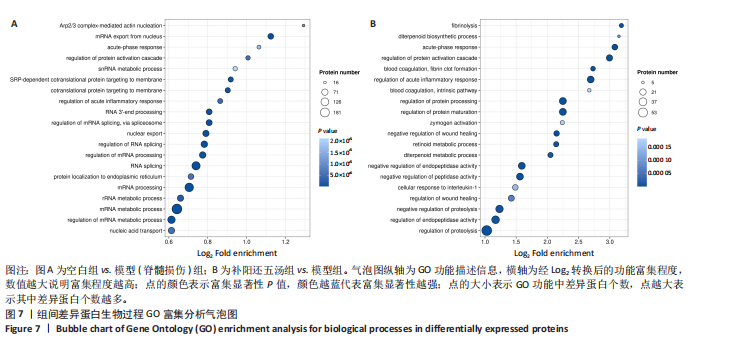
(1)生物过程类差异蛋白GO富集分析:与空白组相比,模型组的差异表达蛋白分析富集到7 988个条目,主要涉及核酸转运mRNA代谢过程的调控、mRNA代谢过程、rRNA代谢过程、加工、蛋白质在内质网的定位、RNA剪接、mRNA加工的调控、RNA剪接的调控、核出口、通过剪接体调控mRNA剪接、RNA3’-末端加工、急性炎症反应的调节、靶向膜的共翻译蛋白、靶向膜的SRP依赖性共翻译蛋白、snRNA代谢过程蛋白质激活级联的调控、急性期反应、输出核、Arp2/3复合物介导的肌动蛋白成核等过程,前20条生物过程类差异蛋白的气泡图,见图7A;与模型组相比,补阳还五汤组的差异表达蛋白生物过程分析富集到3 571个生物过程条目,主要存在于蛋白水解调节内肽酶活性的调节、蛋白水解的负调控、伤口愈合的调节、白细胞介素1的细胞反应、肽酶活性的负调控、内肽酶活性的负调控、二萜代谢过程、维甲酸代谢过程、伤口愈合的负调控、酶原激活、蛋白质加工水平的调控、蛋白质加工的调控、凝血,内在途径、急性炎症反应的调节、凝血、纤维蛋白凝块形成、蛋白质激活级联的调控、急性期反应、二萜生物合成过程、纤维蛋白溶解等生物过程,前20条生物过程的气泡图,见图7B。
总之,两个对比组之间差异蛋白均富集到急性期反应、蛋白质激活级联的调节过程的调节、急性炎症反应的调节等生物过程。
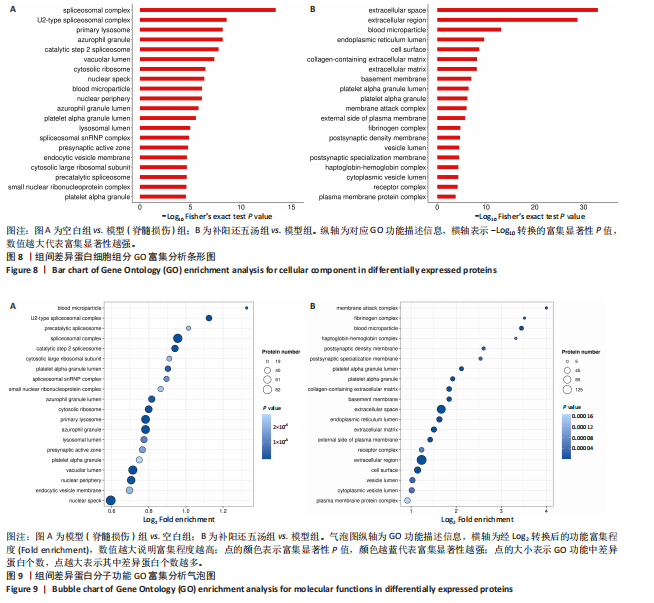
(2)细胞组分类GO注释中分析:模型组的差异表达蛋白与空白组相比,一共富集到984个条目,主要涉及血小板α颗粒、小核核糖核蛋白复合物、预催化剪接体、细胞质大核糖体亚基、内吞囊泡膜、突触前活性区、剪接体snRNP复合体、溶酶体腔、血小板α颗粒管腔、嗜天青颗粒管腔、核外围、血液微粒、核斑点、细胞质核糖体、囊泡腔、催化步骤2剪接体、嗜苯胺蓝粒、初级溶酶体、U2型剪接体复合体、剪接体复合体等细胞组分,前20条细胞组分的条形图,见图8A;补阳还五汤组的差异表达蛋白与模型组相比,总共富集到414个细胞组分类别条目,主要存在于质膜蛋白复合物、受体复合物、细胞质囊腔、触珠蛋白-血红蛋白复合物、突触后特化膜、囊泡腔、突触后密度膜、纤维蛋白原复合物、质膜外侧、膜攻击复合物、血小板α颗粒、血小板α颗粒管腔、基底膜、细胞外基质、含胶原蛋白的细胞外基质、细胞表面、内质网内腔、血液微粒、胞外区、细胞外空间等细胞组分,前20条细胞组分的条形图,见图8B。
总之,两个对比组之间差异蛋白均富集到血小板α颗粒、血液微粒、囊泡腔等细胞组分。
(3)分子功能GO注释中分析:与空白组相比,模型组的差异表达蛋白富集到7 988个条目,主要涉及组蛋白结合、维生素结合、水解酶活性,作用于糖基键、水解酶活性,水解O-糖基化合物、抗原结合、前mRNA结合、启动子特异性染色质结合、丝氨酸型内肽酶抑制剂活性、类异戊二烯结合、补体结合、维甲酸结合、核孔的结构组成、免疫球蛋白结合、免疫球蛋白受体结合、细胞周期蛋白依赖性蛋白激酶活性、细胞周期蛋白依赖性蛋白丝氨酸/苏氨酸激酶活性、己糖胺酶活性、配体门控单原子阴离子通道活性、甘露糖基寡糖甘露糖苷酶活性、二肽酶活性等生物功能,前20条分子功能的气泡图,见图9A;与模型组相比,补阳还五汤组的差异表达蛋白富集到3 571个分子功能条目,主要存在于信号受体结合、阳离子结合、金属离子结合、肽酶活性、内肽酶活性、钙离子结合、酶抑制剂活性、糖胺聚糖结合、胶原结合、肽酶调节活性、丝氨酸水解酶活性、丝氨酸型肽酶活性、内肽酶调节活性、肽酶抑制剂活性、内肽酶抑制剂活性、丝氨酸型内肽酶活性、谷氨酸受体活性、发射机门控信道活动、发射器门控单原子离子通道活动、丝氨酸型内肽酶抑制剂活性等生物功能,前20条分子功能的气泡图,见图9B。
总之,两个对比组之间差异蛋白均富集到丝氨酸型内肽酶抑制剂活性等生物功能。
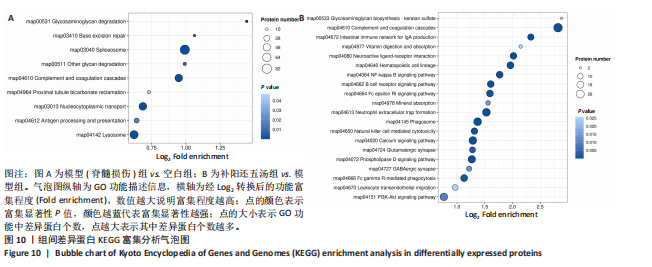
2.4.6 差异蛋白KEGG富集通路分析 通过对空白组与模型组之间的差异蛋白进行富集分析,富集到的KEGG信号通路共9条,主要涉及map04142溶酶体、map04612抗原处理和呈递、map03013核质运输、map04964近端小管碳酸氢盐回收、map04610补体和凝血级联、ap00511其他聚糖降解、地图03040拼接体、map03410基底切除修复、map00531糖胺聚糖降解等信号通路,前9条KEGG的气泡图,见图10A;通过对模型组与补阳还五汤组之间的差异蛋白进行富集分析,富集到的KEGG信号通路共26条,主要涉及map04151PI3K?Akt信号通路、map04670白细胞经内皮细胞迁移、map04666FcγR介导的吞噬作用、map04727GABA能突触、map04072磷脂酶D信号通路、map04724谷氨酸能突触、map04020钙信号通路、map04650自然杀伤细胞介导的细胞毒性、map04145幻影体、map04613中性粒细胞胞外陷阱形成、map04978矿物吸收、map04664FcεRI信号通路、map04662B细胞受体信号通路、map04064NF-κB信号通路、map04640造血细胞谱系、map04080神经活性配体-受体相互作用、map04977维生素消化和吸收、map04672用于IgA生成的肠道免疫网络、map04610补体和凝血级联、map00533糖胺聚糖生物合成-硫酸角蛋白等信号通路,前20条KEGG的气泡图,见图10B。
总之,两个对比组之间差异蛋白共同富集到map04610补体和凝血级联信号,该通路可能参与了补阳还五汤对急性急性脊髓损伤治疗过程。
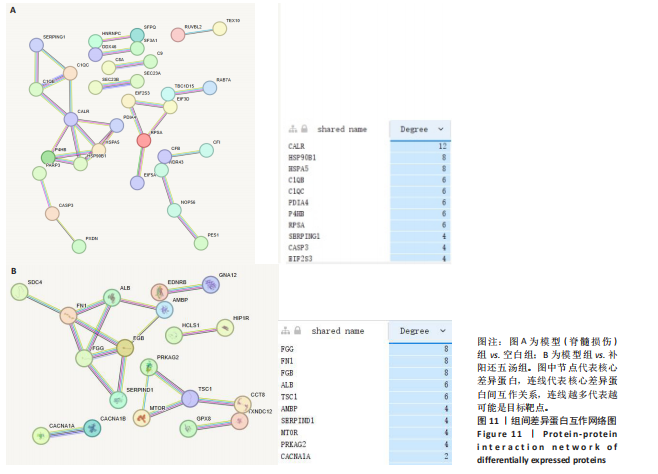
2.4.7 蛋白互作网络分析 模型组与空白组相比,选择差异系数最大的前100位差异蛋白进行PPI分析中发现共同差异蛋白Degree排名10前位有CALR、HSP90B1、HSPA5、C1QB、C1QC、PDIA4、P4HB、RPSA、SERPING1、CASP3等共10种蛋白位于功能网络节点,见图11A。
模型组与空白组相比,选择差异系数最大的前100位差异蛋白进行PPI分析中发现共同差异蛋白Degree排名10前位有FGG、FN1、FGB、ALB、TSC1、AMBP、SERPIND1、MTOR、PRKAG2、CACNA1A等共10种蛋白位于功能网络节点,见图11B。
2.5 转录组学分析结果
2.5.1 测序数据质量与比对分析 转录组测序共获得16组样本,原始数据(RawReads)总量为4.8×107-6.5×107条。过滤碱基平均质量值< 20和无法确定的碱基的数据后,有效数据(CleanReads)为4.7×107-6.5×107条,碱基质量值Q30≥92.00%,各个样本中的表达量FPKM值接近,转录本的丰度信息较高,说明各个样本基因表达水平较高,测序数据质量可靠。样本间相关系数均> 0.8,表示样本选择合理,样本间相关性、实验可靠性较高。
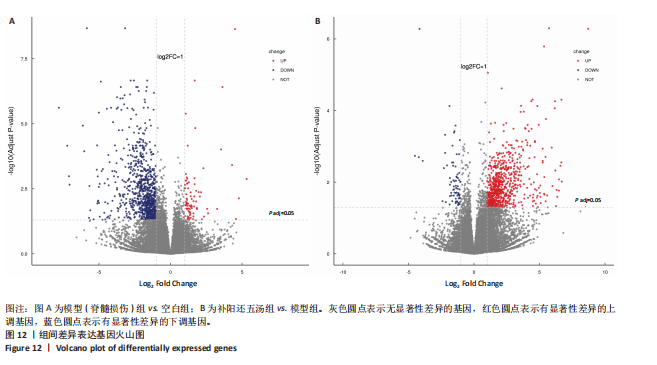
2.5.2 差异表达基因分析 在空白组与模型组的比较中,共识别出968个差异表达基因,其中84个差异表达基因表达上调,884个差异表达基因表达下调(见图12A)。在补阳还五汤组与模型组的比较中,共发现801个差异表达基因,其中727个差异表达基因表达上调,74个差异表达基因表达下调(见图12B)。在不同组的两两比较中,共有593个差异表达基因发生变化,其中24个差异表达基因因无法注释而被排除在分析之外。在差异表达基因表达回调方面,模型组中上调的727个差异表达基因经过补阳还五汤干预后,有524个表达发生逆转回调;模型组中下调的74个差异表达基因经过干预后,有45个基因表达发生逆转回调。
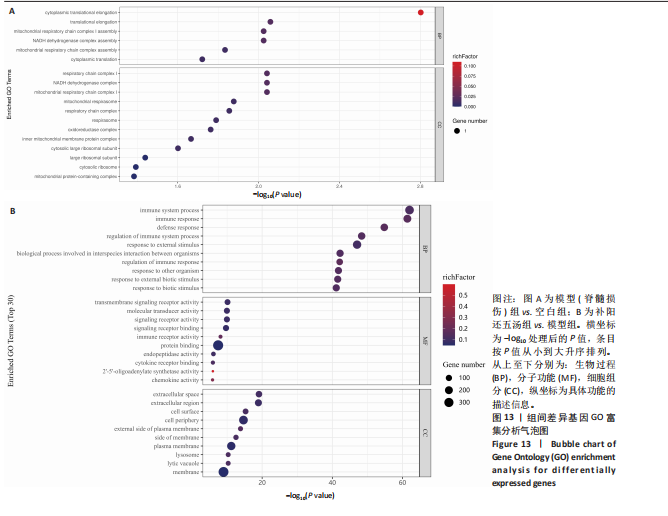
2.5.3 GO功能富集分析 GO功能分类可分为生物进程、细胞组分、生物功能等3个方面。通过对空白组与模型组之间的差异基因进行GO功能富集分析,富集到的9 604个条目;与模型组相比,补阳还五汤组的差异表达基因进行GO功能富集分析,富集到7 963个条目。
(1) GO注释中生物过程分析:与空白组相比,模型组的差异表达基因GO富集,主要涉及细胞质翻译,线粒体呼吸链复合物组装,NADH脱氢酶复合物组装,线粒体呼吸链复合物Ⅰ组装,平移伸长率,细胞质平移伸长等过程,见图13A;与模型组相比,补阳还五汤组的差异表达基因GO富集,主要存在于对生物刺激的反应、外部生物刺激的反应、对其他微生物的反应、疫反应的调节、生物体之间种间相互作用的生物过程、对外部刺激的反应、免疫系统过程的调节防御响应、免疫应答免疫系统过程等生物过程,差异基因生物过程的气泡图,见图13B。
(2) GO注释中细胞组分分析:与空白组相比,模型组的差异表达基因GO富集,主要涉及含线粒体蛋白的复合物、胞质核糖体、大核糖体亚基、胞质大核糖体亚基、线粒体内膜蛋白复合物、氧化还原酶复合物、呼吸体、呼吸链复合体、线粒体呼吸体、线粒体呼吸链复合物Ⅰ、NADH脱氢酶复合物、呼吸链复合物Ⅰ细胞组分,见图13A;与模型组相比,补阳还五汤组的差异表达基因GO富集,主要存在于膜、溶解液泡、溶酶体、质膜、膜的侧面、质膜外侧、细胞外围、单元表面、细胞外区域、细胞外间隙等细胞组分,差异基因细胞组分的条形图,见图13B。
(3) GO注释中分子功能分析:与模型组相比,补阳还五汤组的差异表达基因GO富集,主要存在于趋化因子活性、2’-5’-寡腺苷酸合成酶活性、细胞因子受体结合、内肽酶活性、蛋白结合、免疫受体活性、信号受体结合、信号受体活性、分子换能器活性、跨膜信号转导受体活性等生物功能,差异基因分子功能的气泡图,见图13B。
2.5.4 KEGG通路分析
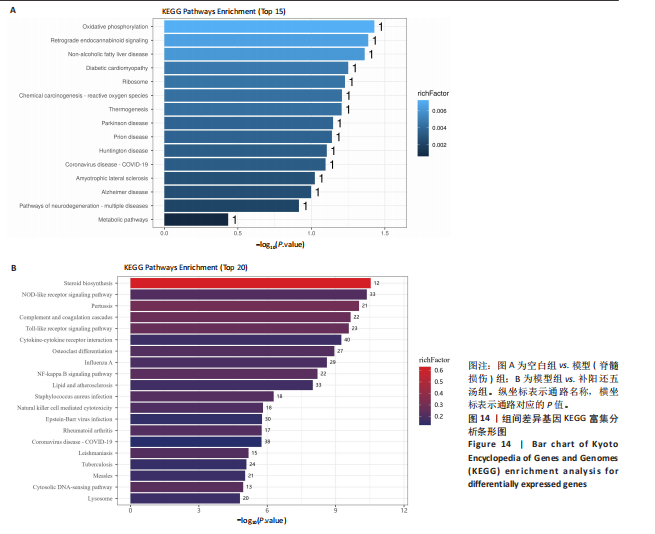
(1)对空白组与模型组之间的KEGG富集分析:一共富集到15条信号通路,主要涉及代谢途径、神经退行性疾病的途径-多种疾病、阿尔茨海默病、肌萎缩侧索硬化症、冠状病毒病?COVID?19、亨廷顿病、朊病毒病、帕金森病、生热作用、化学致癌作用?活性氧、核糖体、糖尿病性心肌病、非酒精性脂肪肝、逆行性内源性大麻素信号传导、氧化磷酸化等15条信号通路,前15条KEGG的条形图见图14A。
(2)对模型组与补阳还五汤组之间的KEGG富集分析:一共富集到298条信号通路,主要涉及溶酶体、胞质DNA感应通路、麻疹、结核、利什曼病、冠状病毒病?COVID?19、类风湿性关节炎、Epstein-Barr病毒感染、自然杀伤细胞介导的细胞毒性、金黄色葡萄球菌感染、脂质和动脉粥样硬化、NF-κB信号通路、甲型流感、破骨细胞分化、细胞因子-细胞因子受体相互作用、Toll样受体信号通路、补体和凝血级联反应、百日咳、NOD样受体信号通路、类固醇生物合成等298条信号通路,前20条KEGG的条形图见图14B。
总之,两个对比组之间差异基因前15条通路中,共同富集到冠状病毒病-COVID-19通路,该通路可能参与了补阳还五汤对急性急性脊髓损伤治疗过程。
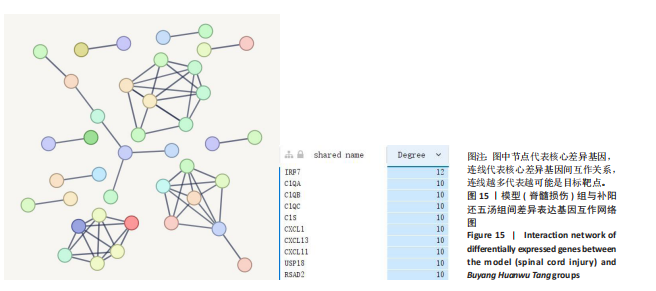
2.5.5 网络核心基因分析 补阳还五汤组上调的45个、下调的524个差异表达基因放到STRING数据库中构建PPI网络,并利用参考物种的互作关系构建互作网络。按照互作score值从高到低排序,选取得分排名Top40的差异基因的互作关系结果,绘制互作网络图。结果显示补阳还五汤中IRF7、C1QA、C1QB、C1QC、C1S、CXCL1、CXCL13、CXCL11、USP18、RSAD2基因是网络核心基因,见图15。
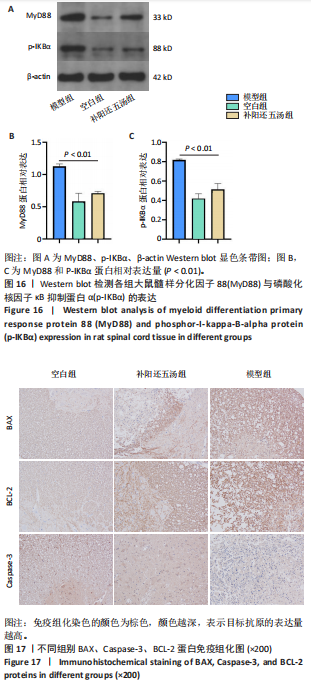
2.5.6 Western blot检测 从差异蛋白中选取髓样分化因子88与磷酸化核因子κB抑制蛋白α(p-IKBα)进行Western blot验证。其中与空白组相比,补阳还五汤组髓样分化因子88与磷酸化核因子κB抑制蛋白α蛋白表达水平略高于空白组(P < 0.01);但与模型组相比,髓样分化因子88与磷酸化核因子κB抑制蛋白α蛋白均远远低于模型组(P < 0.01),见图16,表明补阳还五汤能够抑制髓样分化因子88与磷酸化核因子κB抑制蛋白α蛋白在大鼠急性脊髓损伤中的表达水平。
2.5.7 免疫组化检测 从差异蛋白中选取BAX、Caspase-3、BCL-2进行免疫组化检测验证,如图17所示。补阳还五汤组的BAX、Caspase-3蛋白表达高于空白组,但远低于模型组;补阳还五汤
组的BCL-2蛋白表达低于空白组,但高于模型组;表明补阳还五汤能够抑制BAX、Caspase-3蛋白,促进BCL-2蛋白在大鼠急性脊髓损伤中的表达,抑制细胞凋亡。
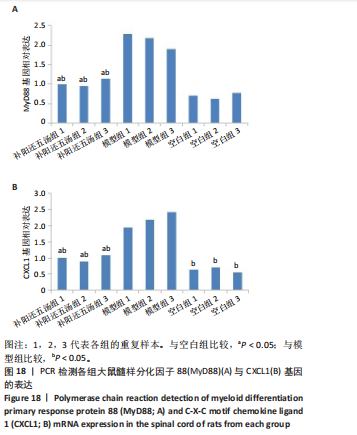
2.5.8 RT-PCR检测 从差异基因中选取髓样分化因子88与CXCL1进行荧光定量PCR检测。其中补阳还五汤组髓样分化因子88表达水平略高于空白组,但都远远低于模型组,见图18A,表明补阳还五汤能够抑制髓样分化因子88在大鼠急性脊髓损伤中的表达水平;补阳还五汤组CXCL1表达水平均高于空白组,但都低于模型组,见图18B,表明补阳还五汤能够抑制CXCL1在大鼠急性脊髓损伤中的表达水平。