中国组织工程研究 ›› 2014, Vol. 18 ›› Issue (51): 8261-8268.doi: 10.3969/j.issn.2095-4344.2014.51.012
• 组织构建与生物活性因子 tissue construction and bioactive factors • 上一篇 下一篇
转化生长因子β1诱导人肾小管上皮细胞转分化:Notch1受体阻断剂的影响
曾 虹,周 懿,姚国媛
- 泸州市人民医院肾病科,四川省泸州市 646000
Transforming growth factor beta 1 induces the transdifferentiation of human renal tubular epithelial cells: the influence of Notch1 receptor specific inhibitor
Zeng Hong, Zhou Yi, Yao Guo-yuan
- Department of Nephrology, Luzhou People’s Hospital, Luzhou 646000, Sichuan Province, China
摘要:
背景:研究发现,在病理状态下各种细胞可通过转化生长因子β1的介导,促进肾小管上皮细胞-间充质细胞转分化为肌成纤维细胞,进而加速肾小管间质纤维化的进展。
目的:验证Notch1受体特异性阻断剂γ-分泌酶抑制剂DAPT能否有效阻断、部分逆转或者完全逆转转化生长因子β1诱导的人肾小管上皮细胞转分化。
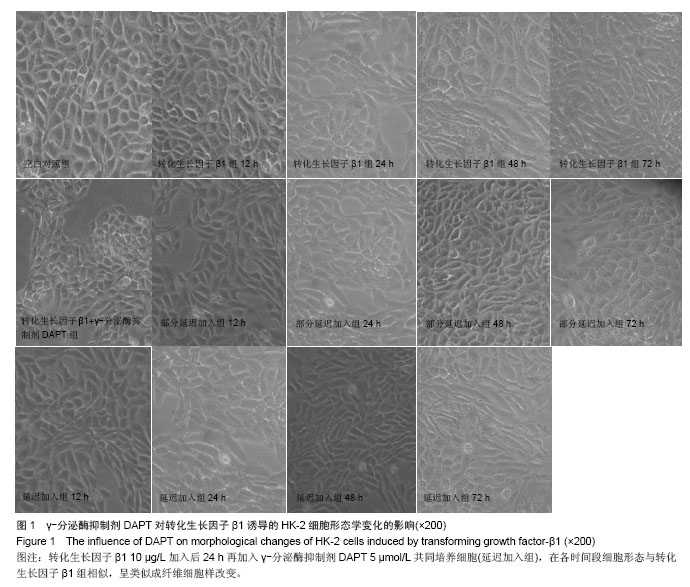
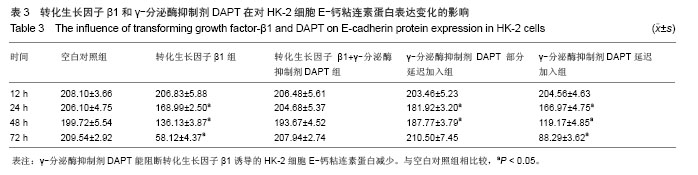
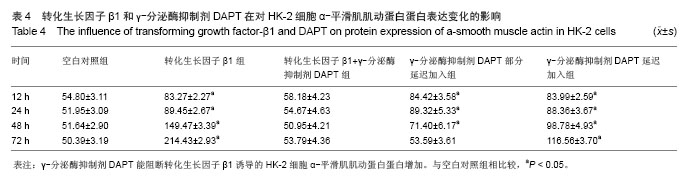
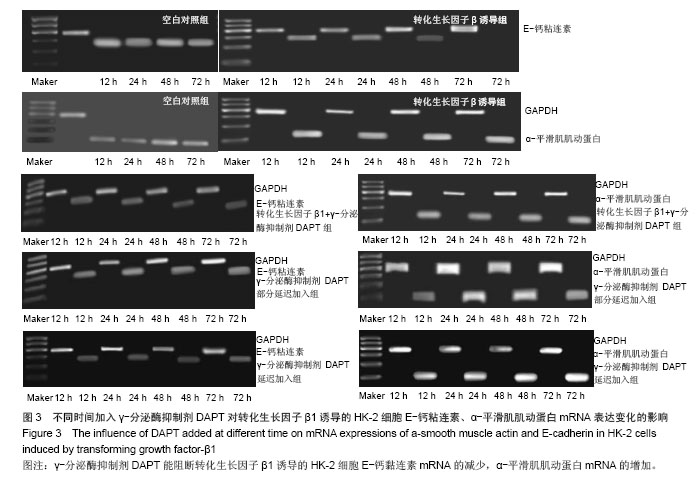
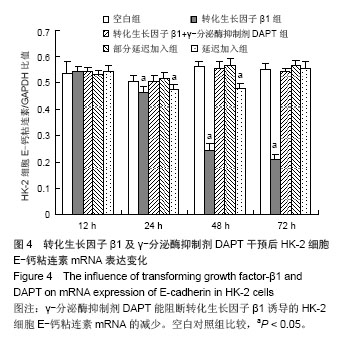
方法:以体外培养的人近端肾小管上皮细胞株HK-2细胞为观察对象,建立转化生长因子β1诱导的肾小管上皮细胞-间充质细胞转分化体外模型,实验分为空白对照组、10 μg/L转化生长因子β1组、转化生长因子β1 (10 μg/L)+ γ-分泌酶抑制剂DAPT(5 μmol/L)组、转化生长因子β1(10 μg/L)+ γ-分泌酶抑制剂DAPT(5 μmol/L)部分延迟加入组、转化生长因子β1 (10 μg/L)+ γ-分泌酶抑制剂DAPT(5 μmol/L)延迟加入组。分别于12,24,48,72 h,在倒置相差显微镜下观察细胞形态变化;用免疫组织化学法和RT-PCR法检测α-平滑肌肌动蛋白、E-钙粘连素蛋白和mRNA表达变化。
结果与结论: ①在干预后在12,24,48,72 h,与空白对照组相比较,转化生长因子β1组α-平滑肌肌动蛋白蛋白和mRNA表达均明显增加(P < 0.05), E-钙粘连素蛋白和mRNA表达逐渐减少(P < 0.05)。②转化生长因子β1+γ-分泌酶抑制剂DAPT组α-平滑肌肌动蛋白的蛋白和mRNA、E-钙粘连素蛋白和mRNA的表达各时间段均接近空白对照组(P > 0.05)。③γ-分泌酶抑制剂DAPT部分延迟加入组,α-平滑肌肌动蛋白的蛋白和mRNA的表达在12 h增加(P < 0.05),之后其表达逐渐减少(P < 0.05),E-钙粘连素蛋白表达在24 h开始减少(P < 0.05),之后逐渐增加,E-钙粘连素mRNA各时间段都与空白对照组相接近,72 h时α-平滑肌肌动蛋白与E-钙粘连素的蛋白及mRNA表达均与空白对照组无显著差异(P > 0.05)。④与空白对照组相比较,延迟加入组各时间点的α-平滑肌肌动蛋白的蛋白表达逐渐增加(P < 0.05), E-钙粘连素蛋白的表达逐渐减少(P < 0.05),但72 h时E-钙粘连素蛋白表达接近空白对照组。结果表明Notch1受体阻断剂γ-分泌酶抑制剂DAPT能够阻断、部分逆转转化生长因子β1所诱导的肾小管上皮细胞-间充质细胞转分化,但不能完全逆转其所诱导的肾小管上皮细胞-间充质细胞转分化。
中图分类号:


.jpg)
.jpg)
.jpg)