Chinese Journal of Tissue Engineering Research ›› 2018, Vol. 22 ›› Issue (17): 2755-2760.doi: 10.3969/j.issn.2095-4344.0531
Previous Articles Next Articles
Impact of oxidative stress on dental pulp stem cells: a key issue in tissue repair
Chen Jun-ting, Zhang Jing-ying
- Medical School of Dalian University, Dalian 116622, Liaoning Province, China
-
Revised:2018-02-23Online:2018-06-18Published:2018-06-18 -
Contact:Zhang Jing-ying, M.D., Associate professor, Medical School of Dalian University, Dalian 116622, Liaoning Province, China -
About author:Chen Jun-ting, Master candidate, Medical School of Dalian University, Dalian 116622, Liaoning Province, China -
Supported by:the National Natural Science Foundation of China, No. 81500890; the Natural Science Foundation of Liaoning Province, No. 20170540028
CLC Number:
Cite this article
Chen Jun-ting, Zhang Jing-ying. Impact of oxidative stress on dental pulp stem cells: a key issue in tissue repair[J]. Chinese Journal of Tissue Engineering Research, 2018, 22(17): 2755-2760.
share this article
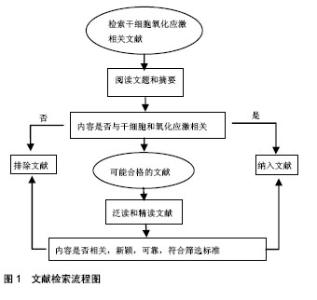
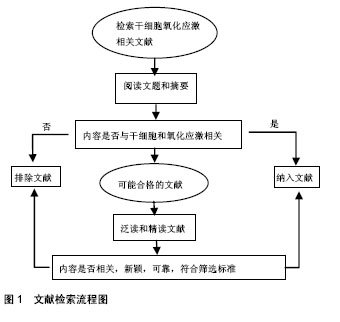
2.1 纳入文献基本情况 纳入文献中的中文文献共19 篇,英文文献共31篇。文献检索流程图见图1。"

2.2 人源牙髓干细胞 2.2.1 人源牙髓干细胞的提出 Gronthos等[8]采用酶消化法在人第三磨牙牙髓组织中分离出一些与骨髓基质干细胞相似的细胞,经诱导可形成分散且高密度的钙化小结,第一次提出了人源牙髓干细胞的概念。人源牙髓干细胞是从牙髓组织中分离出的一种成体干细胞,作为理想的组织工程种子细胞,其具有高度繁殖、多向分化潜能和自我更新能力,对组织工程及再生医学的发展具有重要作用。 2.2.2 人源牙髓干细胞的分子标记 研究发现,在起源组织(颌突上皮和外胚间充质)的微血管周围存在人源牙髓干细胞。Shi等[9]利用间充质干细胞基质细胞抗原能结合牙髓组织血管周细胞抗原的特性来标记人源牙髓干细胞,结果显示10%-20%的STRO-1阳性人源牙髓干细胞被分离出来。 2.2.3 人源牙髓干细胞的生物学特性 高度增殖、自我更新能力和多向分化潜能,是其重要的生物学特性。利用酶消化法分离第三磨牙的牙髓,经鉴定证实为人源牙髓干细胞,且能维持其生物学特性至少25代,随后分别用不同诱导介质在不同的诱导条件下诱导人源牙髓干细胞,发现其具有多向分化的潜能[10]:有研究发现人源牙髓干细胞在进行体外培养时,可以诱导分化成神经元样细胞和神经胶质细胞,出现神经特异质蛋白标志巢蛋白、胶质纤维酸性蛋白等[11];李景辉等[12]从正畸治疗减数拔除的恒前磨牙中提取牙髓组织,酶消化法体外分离培养,单抗Stro-1标记、免疫磁珠阳性分选系统分选获得人源牙髓干细胞,当培养至第3代时,用成脂诱导培养基诱导,油红O染色结果显示为阳性,RT-PCR检测成脂肪向分化相关基因均有阳性表达,证明了人源牙髓干细胞在体外具有成脂细胞分化的潜能。后续有研究报道人恒牙的人源牙髓干细胞在特定诱导条件下的成骨分化潜力[13-14]。Tamaoki等[15]利用病毒载体把c-Myc、Klf4、Oct4、Sox2 和Lin28、Nanog、Oct4、Sox2 因子导入人源牙髓干细胞中,成功获得了多潜能性干细胞。 2.2.4 人源牙髓干细胞体外培养 何飞等[16]通过酶消化法获取第三磨牙来源的人源牙髓干细胞,体外培养计算细胞克隆形成率、免疫组化、RT-PCR法检测细胞表面分子表达,流式细胞仪测定细胞周期,体外诱导检测细胞多向分化能力。结果发现,细胞体积较小,多为成纤维样细胞;免疫组化及RT-PCR法显示细胞有Vimentin、Ⅰ型胶原、GFAP、Nestin和Osteocalcin表达,ALP化学染色阳性,无MyoD和DSPP表达;流式细胞仪检测大多数细胞处于静止期。 2.2.5 人源牙髓干细胞的免疫调节作用 人源牙髓干细胞的免疫调节功能是通过副分泌机制的可溶性因子和细胞因子,通过抑制T淋巴细胞和调节性T细胞上调导致免疫耐受。Zhao等[17]发现Fas配体可以调控人源牙髓干细胞的免疫耐受。人源牙髓干细胞中存在Fas配体,当Fas配体敲除后,能降低诱导T细胞凋亡的能力。这表明Fas配体可以调控人源牙髓干细胞的免疫能力。 2.2.6 人源牙髓干细胞的研究意义 人源牙髓干细胞具有以下优点[18]:①来源丰富,易采集。6-11岁的小孩自然脱落的牙齿、正畸减数拔牙、以及成人需要拔除的智齿中都含有丰富的人源牙髓干细胞;②不良反应少,具有免疫抗炎作用,可用于异体移植研究。免疫原性低,无须经过严格配对,不会引起强烈的排斥反应,具有一定的免疫调节功能;③一定条件下可诱导分化成釉质和牙本质;④可以体外分离扩增;⑤无伦理争议。人源牙髓干细胞在口腔应用方面,作为种子细胞它可以用于再生牙根、骨、牙周软硬组织,可广泛应用于牙周病治疗、牙周再生、颌骨再生、牙齿再生、口腔颌面部放射性骨坏死,双膦酸盐造成的颌骨坏死等疾病,甚至使牙齿再生成为指日可待的新希望。另外,随着越来越多的深入研究发现,乳牙人源牙髓干细胞在治疗阿尔茨海默病、帕金森病、急性炎症、糖尿病等动物实验模型中发挥显著作用[19]。 2.3 人源牙髓干细胞在组织工程中的应用 2.3.1 人源牙髓干细胞修复骨缺损 在培养人源牙髓干细胞过程中加入含有地塞米松、抗坏血酸、β-甘油磷酸钠的成骨诱导液,在其诱导作用下人源牙髓干细胞可向成骨细胞方向分化,且能表达成骨标志物。Ito等[20]将富含血小板的血浆与人源牙髓干细胞的混合物植入犬上颌骨缺损中,8周后植入种植体,然后再培养8周,结果发现种植体周围不仅有骨组织形成,而且提高了种植体与周围骨组织的结合能力。有学者将新西兰兔随机分为实验组和对照组,分别在下颌无牙区制造10 mm×4 mm×4 mm的骨缺损,空白组加入浸有PBS的Bio-oss骨粉,实验组加入人源牙髓干细胞和Bio-oss骨粉,术后6周处死。实验发现,空白组纤维结缔组织较疏松,苏木精-伊红染色未见明显新骨形成,而实验组骨缺损区愈合良好,可见新生骨形成,并且与周围骨结合良好[21]。从而证实了人源牙髓干细胞具有较好的成骨能力。 2.3.2 人源牙髓干细胞在再生牙组织工程中的应用Iohara等[22]将犬牙中获得的牙髓侧群细胞自体移植入活髓切断模型中,14 d内可形成含毛细血管和神经细胞的完整牙髓。因此将人源牙髓干细胞与可降解支架材料复合,形成牙本质-牙髓复合体修复受损牙髓,成为组织工程修复受损组织器官一种安全有效的治疗手段。 2.3.3 人源牙髓干细胞成肌肉分化 研究显示,人源牙髓干细胞可表达典型的成肌细胞标志物α-平滑肌肌动蛋白[23]。另外,注射人源牙髓干细胞到心肌梗死动物模型内能有效恢复部分心肌功能,减小心肌梗死面积,这提示对人源牙髓干细胞的深入研究可有助于心肌梗死类疾病的治疗。 2.3.4 人源牙髓干细胞成神经元分化 多种实验研究结果表明人源牙髓干细胞具有向神经元细胞分化的潜能。王亦菁等[24]通过实验研究,在一定的神经诱导条件下,人源牙髓干细胞采用一定的神经元标志物鉴定发现NSE染色阳性,表明人源牙髓干细胞有向神经元分化的潜能。有学者将分离获得的人源牙髓干细胞经诱导后用于治疗中风的大鼠,发现能够帮助中风后的受损组织恢复功能。孔杰等[26]发现骨髓间充质干细胞对大鼠神经损伤有治疗效应,能上调组织抗氧化能力,促进神经因子分泌,并调节抗氧化物酶活性保护受损神经,减少大鼠缺血性神经元凋亡,从而产生治疗效应。以上实验研究均表明人源牙髓干细胞能修复神经系统损伤组织,为治疗神经系统疾病提供新方法。 2.4 氧化应激 2.4.1 氧化应激的提出及损伤机制 伴随着生物体逐渐衰老,机体抗氧化能力不断下降,导致自由基不断积累,机体在有害刺激因素作用下,氧自由基产生过多或发生代谢障碍,细胞体内氧化及抗氧化体系失衡,诱发基因突变、蛋白质变性和脂质过氧化,进而损伤细胞溶酶体、线粒体等,导致细胞的生理生化及代谢功能障碍。 2.4.2 氧化应激反应与疾病的关系 有资料表明心血管系统疾病如原发性高血压、动脉粥样硬化以及糖尿病患者的发病机制均与氧化应激有关;氧化应激产物激活某些信号通路促进炎性递质释放,与肺部炎症有明确相关性;氧化应激时黄嘌呤氧化酶激活,患者产生的自由基损伤脑细胞,对神经元造成损伤;糖尿病患者普遍存在氧化应激反应增强,与其并发症的发生和发展关系密切[27]。 2.4.3 氧化应激导致的全身性疾病对口腔牙种植成功率的影响 随着口腔种植技术的推广应用,患有糖尿病、牙周病等患者的种植需求越来越多,但是种植牙在该类疾病中应用的成功率相对较低。牙种植术在健康个体中常规应用有较高的成功率,达95%-99%,但是在糖尿病患者中应用的成功率仅为85.5%-94.3%[28]。种植牙在糖尿病患者中的高失败率和较低骨结合率已经得到较多的实验及临床证实[29-30]。 2.5 目前氧化应激对人源牙髓干细胞影响的研究进展 2.5.1 不同浓度过氧化氢诱导人源牙髓干细胞氧化应激损伤 双氧水是一种重要的活性氧,能自由弥散且稳定,可以导致不同形式的氧化改变,触发细胞死亡[31-32]。徐克等[33]将人源牙髓干细胞分为3组:分别用PBS,100 μmol/L H2O2、200 μmol/L H2O2分别刺激2 h,倒置显微镜观察细胞形态,MTT、β-半乳糖苷酶染色观察细胞增殖及衰老程度,免疫荧光检测细胞骨架排列情况和sirt1蛋白表达,测定P16、sirtⅠ蛋白表达。结果显示H2O2能有效诱导人源牙髓干细胞氧化应激损伤,细胞形态由成纤维细胞样梭形外观,体积变大、扁平、失去纺锤样结构。同时引起人源牙髓干细胞增殖能力降低,细胞骨架排列紊乱,细胞衰老增强,在此过程中,激活p16、p53等衰老信号通路的活化,P16表达上调,Sirt1蛋白的表达受到抑制,细胞周期停滞。 2.5.2 氧化应激增强抗氧化防御基因上调 Alami等[34]研究表明环境氧气张力(体积分数21%O2)通常用于体外培养,但体内生理水平范围在3%-6%之间。通过比较21%对3%氧气张力下人源牙髓干细胞的增殖,发现由于氧化应激增强细胞周期调节蛋白P21、P38激活、NRF-2信号通路抗氧化防御基因上调。 2.5.3 人源牙髓干细胞在抗衰老和组织工程领域的潜在应用 人源牙髓干细胞因具有多向分化能力在组织中具有潜在的应用价值,但是在长期的细胞培养和氧化应激过程中,人源牙髓干细胞的组织再生潜力受到长期的负面调节。为了减少人源牙髓干细胞的细胞衰老和氧化应激,超氧化物歧化酶1的细胞内递送系统发展起来。Jung等[35]将超氧化物歧化酶1与称为低分子量细胞穿透肽结合,发现,氧自由基对干细胞影响的主要问题是减少其分化和自我更新潜力。H2O2导致的早衰细胞中p53激活和p21表达可能是由于H2O2导致的DNA氧化损伤。P53是衰老机体的重要标志分子,在细胞衰老和凋亡中起重要作用[36]。另外,还有研究表明p53/p21途径与p16/pRb途径共同诱导早熟性衰老,p53不仅可以上调促凋亡因子,还能下调抗凋亡因子[37-38]。 2.5.4 龋牙髓干细胞具有更高的抗氧化蛋白表达 Ma等[39]通过使用2-D DIGE鉴定人源牙髓干细胞和龋牙髓干细胞差异表达的蛋白质研究结果表明,在4-7的pH值范围内,人源牙髓干细胞和龋牙髓干细胞之间有18个蛋白点差异表达。这些不同表达的蛋白质主要参与调节细胞增殖、分化、细胞骨架和运动。此外,研究结果表明,龋牙髓干细胞具有更高的抗氧化蛋白表达,可以保护龋牙髓干细胞免受氧化应激。此研究对理解人源牙髓干细胞在牙本质-牙髓复合物形成中的分子矿化机制有重大意义。 2.5.5 氧化应激对干细胞重编程的影响 氧化应激、低氧、热处理等均可能引发细胞的重编程过程,使已分化的干细胞去分化和重编程,提供了组织工程种子细胞应用的新途径[40]。当机体发生氧化应激反应时,细胞将从已分化状态变为未分化状态,重新开始细胞周期。Ondre等[41]研究发现氧化应激状态时细胞的去分化和重编程过程与氧自由基失衡的时间及各自的量有关。未来针对氧化应激等微环境的改变对细胞重编程的影响机制的研究将为今后诱导多能干细胞提供理论依据。 2.5.6 多种信号转导途径参与了氧化应激骨髓间充质干细胞的衰老及凋亡 研究表明,AKT、p53、p38MAPK、c-Jun氨基末端激酶等信号分子均参与了氧自由基聚集导致的氧化应激状态下基质干细胞的衰老和凋亡。AKT信号途径磷酸化能激活抗凋亡蛋白,使凋亡因子失活,同时还能增加p38β的活性来降低ROS水平,抑制凋亡分子JNK,是细胞应激状态下存活的重要信号途径[42]。p38MAPK途径可以通过细胞色素C的释放和Bax的转移诱导基质干细胞的凋亡[43]。APE/REF-1酶是一种可以修复氧化性损伤的限速酶,能阻碍氧自由基聚集对机体的损伤,氧自由基聚集可以诱导其表达。研究显示mTOR及其下游蛋白在没有干预措施时活性较低,参与正常情况下的细胞增殖分化,当施加低浓度氧化应激干预措施时,mTOR及其下游蛋白表达增高;高浓度的氧化应激干预措施会使mTOR信号通路活性受到明显抑制[44]。胡明等[45]研究发现过表达miR- 486-5p能促进氧化应激状态下细胞的凋亡,而抑制其抑制物作用时可以抑制氧化应激状态下细胞的凋亡,其作用机制可能与Akt通路有关。这提示氧化应激导致的细胞凋亡是多种信号通路共同参与的结果,要综合分析才能认清氧化应激对牙髓干细胞影响的作用机制(表1)。"

| [1] Collart-Dutilleul PV, Chaubron F, Vos JD, et al. Allogenic banking of dental pulp stem cells for innovative therapeutics. World J Stem Cells. 2015; 7(7): 1010–1021.[2] Rodas-Junco BA, Canul-Chan M, Rojas-Herrera RA, et al. Stem cells from dental pulp: what epigenetics can do with your tooth. Front Physiol. 2017; 8: 999.[3] 张蕾,董小伟,郭凌郧,等. PEP-1-CAT抑制氧化应激条件下大鼠骨髓间充质干细胞的凋亡[J]. 湖北医药学院学报,2012,31(4):285-291.[4] 卢文艺,赵明峰.氧化应激对骨髓间充质干细胞的影响[J].中国医学科学院学报, 2012;34(1):90-94.[5] 肖静雯,王陈飞.N-乙酰氨基葡萄糖转移酶调节O-GlcNAc修饰在牙髓干细胞成骨分化中的作用[J].全科口腔医学杂志:电子版,2016,3(16):11-13.[6] Ratajczak J, Bronckaers A, Dillen Y, et al. The Neurovascular properties of dental stem cells and their importance in dental tissue engineering. Stem Cells Int. 2016; 2016: 9762871.[7] Sonoda S, Yamaza H, Ma L, et al. Interferon-gamma improves impaired dentinogenic and immunosuppressive functions of irreversible pulpitis-derived human dental pulp stem cells. Sci Rep. 2016; 6:19286.[8] Gronthos S, Mankani M, Brahim J, et al. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci. 2000;97:13625-13630.[9] Shi S, Gronthos S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J Bone Miner Res. 2003;18:696-704.[10] 郭红延,吴补领,郭希民,等.大鼠牙髓干细胞的培养和鉴定[J].牙体牙髓牙周病学杂志, 2004,14(5):242-245.[11] 方成志,杨于嘉,姚跃,等.人牙髓干细胞的体外培养及神经样诱导分化[J].中国组织工程研究, 2014,18(23):3723-3726.[12] 李景辉,张方明,张振庭.人恒牙牙髓干细胞分化为脂肪细胞的体外实验研究[J].北京口腔医学,2012,20(3):142-146.[13] D'Aquino R, Graziano A, Sampaolesi M, et al. Human postnatal dental pulp cells co-differentiate into osteoblasts and endotheliocytes: a pivotal synergy leading to adult bone tissue formation. Cell Death Differ.2007;14(6):1162-1171.[14] Otaki S, Ueshima S, Shiraishi K, et al. Mesenchymal progenitor cells in adult human dental pulp and their ability to form bone when transplanted into immunocompromised mice. Cell Biol Int. 2013;31:1191-1197.[15] Tamaoki N, Takahashi K, Tanaka T, et al. Dental pulp cells for induced pluripotent stem cell banking. J Dent Res. 2010;89:773.[16] 何飞,谭颖徽,张纲.人牙髓干细胞的体外培养和鉴定[J].华西口腔医学杂志,2005,23(1):75-78.[17] Zhao Y, Wang L, Jin Y, et al. Fas ligand regulates the immunomodulatory properties of dental pulp stem cells. J Dent Res. 2012;91:948-954.[18] Tatullo M, Marrelli M, Shakesheff KM, et al. Dental pulp stem cells: function, isolation and applications in regenerative medicine. J Tissue Eng Regen Med. 2015;9:1205-1216.[19] Wang J, Wang X, Sun Z, et al. Stem cells from human-exfoliated deciduous teeth can differentiate into dopaminergic neuron-like cells. Stem Cells Dev. 2010;19:1375.[20] Ito K, Yamada Y, Nakamura S, et al. Osteogenic potential of effective bone engineering using dental pulp stem cells, bone marrow stem cells, and periosteal cells for osseointegration of dental implants. Int J Oral Maxillofac Surg. 2011;26:947.[21] 吴中明,木合塔尔•霍加,买布拜木•买买提依明,等.牙髓干细胞在修复牙槽骨缺损的组织学观察[J].石河子大学学报(自科版),2015, 1(33):107-111.[22] Iohara K, Imabayashi K, Ishizaka R, et al. Complete pulp regeneration after pulpectomy by transplantation of CD105+ stem cells with stromal cell-derived factor-1. Tissue Eng Part A. 2011; 17:1911.[23] 王洁,刘加强,房兵.牙髓干细胞在组织工程中的研究进展[J].口腔颌面修复学杂志, 2013,14(3):183-186.[24] 王亦菁,徐莉,金岩,等.体外诱导牙髓干细胞、外胚间充质干细胞向神经分化的研究[J].牙体牙髓牙周病学杂志,2009,19(11):616-618.[25] Leong WK, Henshall TL, Arthur A, et al. Human adult dental pulp stem cells enhance poststroke functional recovery through non-neural replacement mechanisms. Stem Cells Transl Med. 2012;1:177.[26] 孔杰,牛朝诗,杨艳艳,等.骨髓间质干细胞移植对大鼠颅脑损伤后氧化应激的影响[J].立体定向和功能性神经外科杂志,2006,19(3): 149-152.[27] 王全伟,凡文博,王智昊,等.氧化应激与心血管疾病关系的研究进展[J].中国老年学, 2014,34(1):192-194.[28] 邹桂克. bFGF局部应用对2型糖尿病大鼠种植体骨结合的影响[D].第四军医大学,2011.[29] Nevins ML, Karimbux NY, Weber HP, et al. Wound healing around endosseous implants in experimental diabetes. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 1998;87:293.[30] Mccracken M, Lemons JE, Rahemtulla F, et al. Bone response to titanium alloy implants placed in diabetic rats. Int J Oral Maxillofac Implants. 2000;15:345.[31] Jian Z, Li K, Liu L, et al. Heme oxygenase-1 protects human melanocytes from H2O2-induced oxidative stress via the Nrf2-ARE pathway. J Invest Dermatol. 2011;131:1420.[32] Kim JH, Choi W, Lee JH, et al. Astaxanthin Inhibits H_2O_2-Mediated Apoptotic Cell Death in Mouse Neural Progenitor Cells via Modulation of P38 and MEK Signaling Pathways. J Microbiol Biotechnol. 2009;19:1355-1363.[33] 徐克,冯桂娟,冯兴梅,等.过氧化氢刺激人牙髓干细胞的衰老进程[J].中国组织工程研究, 2016,20(10):1481-1487.[34] Alami M, Viña-Almunia J, Gambini J, et al. Activation of p38, p21, and NRF-2 mediates decreased proliferation of human dental pulp stem cells cultured under 21% O2. Stem Cell Reports. 2014;3:566-573.[35] Jung CY, Yeon LJ, Pyoung CC, et al. Cell-penetrating superoxide dismutase attenuates oxidative stress-induced senescence by regulating the p53-p21Cip1pathway and restores osteoblastic differentiation in human dental pulp stem cells.Int J Nanomedicine. 2012;7:5091-5106.[36] Levine AJ. p53, the cellular gatekeeper for growth and division. Cell. 1997;88:323-331.[37] Yang DG, Liu L, Zheng XY. Cyclin-dependent kinase inhibitor p16 INK4a and telomerase may co-modulate endothelial progenitor cells senescence. Ageing Res Rev. 2008;7:137-146.[38] Mimeault M, Batra SK. Recent insights into the molecular mechanisms involved in aging and the malignant transformation of adult stem/progenitor cells and their therapeutic implications. Ageing Res Rev. 2009;8:94-112.[39] Ma D, Cui L, Gao J, et al. Proteomic analysis of mesenchymal stem cells from normal and deep carious dental pulp. PLoS One. 2014;9:e97026.[40] 凌均棨,彭正军,刘路.细胞重编程及其影响因素[J].国际口腔医学杂志,2014,41(3):300-303.[41] Ond?ej V, Protivánková I, Piterková J, et al. Recondensation level of repetitive sequences in the plant protoplast nucleus is limited by oxidative stress. J Exp Bot.2010;61:2395.[42] Zhang W, Su X, Gao Y, et al. Berberine protects mesenchymal stem cells against hypoxia-induced apoptosis in vitro. Biol Pharm Bull. 2009;32:1335-1342.[43] Wei H, Li Z, Hu S, et al. Apoptosis of mesenchymal stem cells induced by hydrogen peroxide concerns both endoplasmic reticulum stress and mitochondrial death pathway through regulation of caspases, p38 and JNK. J Cell Biochem. 2010;111: 967-978.[44] 黄文秋,黄宏,徐祥,等.mTOR及其下游信号通路在骨髓间充质干细胞氧化应激损伤中的变化及作用[J].第三军医大学学报,2013,35(2): 114-118.[45] 胡明,黎佼,刘宁宁,等.miR-486-5p在氧化应激引起人骨髓间充质干细胞凋亡中的作用[J]. 中国病理生理杂志,2015,31(3):524-529.[46] 束波,刘志江,范芳.基质细胞衍生因子1预处理抑制大鼠骨髓间充质干细胞的凋亡[J].中国组织工程研究,2012,16(32): 5903-5908.[47] Dai Z, Li Y, Quarles LD, et al. Resveratrol enhances proliferation and osteoblastic differentiation in human mesenchymal stem cells via ER-dependent ERK1/2 activation. Phytomedicine. 2007;14: 806-814.[48] Denu RA, Hematti P. Effects of oxidative stress on mesenchymal stem cell biology. Oxid Med Cell Longev.2016;2016:1-9.[49] Volonte D, Zou H, Bartholomew JN, et al. Oxidative stress-induced inhibition of sirt1 by caveolin-1 promotes p53-dependent premature senescence and stimulates the secretion of interleukin 6 (IL-6). J Biol Chem. 2015;290:4202.[50] Kao CL, Chen LK, Chang YL, et al. Resveratrol protects human endothelium from H(2)O(2)-induced oxidative stress and senescence via SirT1 activation. J Thromb Thrombolysis. 2010; 17:970. |
| [1] | Pu Rui, Chen Ziyang, Yuan Lingyan. Characteristics and effects of exosomes from different cell sources in cardioprotection [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(在线): 1-. |
| [2] | Lin Qingfan, Xie Yixin, Chen Wanqing, Ye Zhenzhong, Chen Youfang. Human placenta-derived mesenchymal stem cell conditioned medium can upregulate BeWo cell viability and zonula occludens expression under hypoxia [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(在线): 4970-4975. |
| [3] | Zhang Tongtong, Wang Zhonghua, Wen Jie, Song Yuxin, Liu Lin. Application of three-dimensional printing model in surgical resection and reconstruction of cervical tumor [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(9): 1335-1339. |
| [4] | Zhang Xiumei, Zhai Yunkai, Zhao Jie, Zhao Meng. Research hotspots of organoid models in recent 10 years: a search in domestic and foreign databases [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(8): 1249-1255. |
| [5] | Hou Jingying, Yu Menglei, Guo Tianzhu, Long Huibao, Wu Hao. Hypoxia preconditioning promotes bone marrow mesenchymal stem cells survival and vascularization through the activation of HIF-1α/MALAT1/VEGFA pathway [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 985-990. |
| [6] | Shi Yangyang, Qin Yingfei, Wu Fuling, He Xiao, Zhang Xuejing. Pretreatment of placental mesenchymal stem cells to prevent bronchiolitis in mice [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 991-995. |
| [7] | Liang Xueqi, Guo Lijiao, Chen Hejie, Wu Jie, Sun Yaqi, Xing Zhikun, Zou Hailiang, Chen Xueling, Wu Xiangwei. Alveolar echinococcosis protoscolices inhibits the differentiation of bone marrow mesenchymal stem cells into fibroblasts [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 996-1001. |
| [8] | Fan Quanbao, Luo Huina, Wang Bingyun, Chen Shengfeng, Cui Lianxu, Jiang Wenkang, Zhao Mingming, Wang Jingjing, Luo Dongzhang, Chen Zhisheng, Bai Yinshan, Liu Canying, Zhang Hui. Biological characteristics of canine adipose-derived mesenchymal stem cells cultured in hypoxia [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1002-1007. |
| [9] | Geng Yao, Yin Zhiliang, Li Xingping, Xiao Dongqin, Hou Weiguang. Role of hsa-miRNA-223-3p in regulating osteogenic differentiation of human bone marrow mesenchymal stem cells [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1008-1013. |
| [10] | Lun Zhigang, Jin Jing, Wang Tianyan, Li Aimin. Effect of peroxiredoxin 6 on proliferation and differentiation of bone marrow mesenchymal stem cells into neural lineage in vitro [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1014-1018. |
| [11] | Zhu Xuefen, Huang Cheng, Ding Jian, Dai Yongping, Liu Yuanbing, Le Lixiang, Wang Liangliang, Yang Jiandong. Mechanism of bone marrow mesenchymal stem cells differentiation into functional neurons induced by glial cell line derived neurotrophic factor [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1019-1025. |
| [12] | Duan Liyun, Cao Xiaocang. Human placenta mesenchymal stem cells-derived extracellular vesicles regulate collagen deposition in intestinal mucosa of mice with colitis [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1026-1031. |
| [13] | Pei Lili, Sun Guicai, Wang Di. Salvianolic acid B inhibits oxidative damage of bone marrow mesenchymal stem cells and promotes differentiation into cardiomyocytes [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1032-1036. |
| [14] | Guan Qian, Luan Zuo, Ye Dou, Yang Yinxiang, Wang Zhaoyan, Wang Qian, Yao Ruiqin. Morphological changes in human oligodendrocyte progenitor cells during passage [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1045-1049. |
| [15] | Wang Zhengdong, Huang Na, Chen Jingxian, Zheng Zuobing, Hu Xinyu, Li Mei, Su Xiao, Su Xuesen, Yan Nan. Inhibitory effects of sodium butyrate on microglial activation and expression of inflammatory factors induced by fluorosis [J]. Chinese Journal of Tissue Engineering Research, 2021, 25(7): 1075-1080. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||