Chinese Journal of Tissue Engineering Research ›› 2026, Vol. 30 ›› Issue (22): 5792-5803.doi: 10.12307/2026.180
Previous Articles Next Articles
Strategies for the application of miRNA-targeted therapy in the treatment of osteoporosis
Wu Lingjie1, Zheng Kaiyuan2, Wang Guangrong1, Yin Chong1, 3
- 1Department of Laboratory Medicine, 2Department of Rehabilitation, Affiliated Hospital of North Sichuan Medical College, Nanchong 637000, Sichuan Province, China; 3Translational Medicine Research Center, School of Laboratory Medicine, North Sichuan Medical College, Nanchong 637000, Sichuan Province, China
-
Received:2025-06-16Accepted:2025-08-31Online:2026-08-08Published:2025-12-27 -
Contact:Yin Chong, PhD, Associate researcher, Department of Laboratory Medicine, Affiliated Hospital of North Sichuan Medical College, Nanchong 637000, Sichuan Province, China; Translational Medicine Research Center, School of Laboratory Medicine, North Sichuan Medical College, Nanchong 637000, Sichuan Province, China -
About author:Wu Lingjie, MS candidate, Department of Laboratory Medicine, Affiliated Hospital of North Sichuan Medical College, Nanchong 637000, Sichuan Province, China -
Supported by:National Natural Science Foundation of China (Youth Project), No. 32000924 (to YC); Sichuan Provincial Natural Science Foundation (Youth Project), No. 23NSFSC6012 (to YC); 2022 Best Candidate Project of North Sichuan Medical College Affiliated Hospital, No. 2022JB007 (to YC)
CLC Number:
Cite this article
Wu Lingjie, Zheng Kaiyuan, Wang Guangrong, Yin Chong . Strategies for the application of miRNA-targeted therapy in the treatment of osteoporosis[J]. Chinese Journal of Tissue Engineering Research, 2026, 30(22): 5792-5803.
share this article
Add to citation manager EndNote|Reference Manager|ProCite|BibTeX|RefWorks
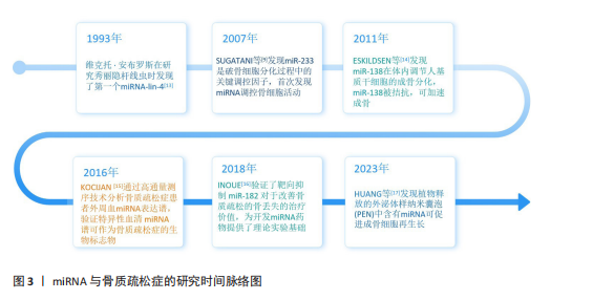
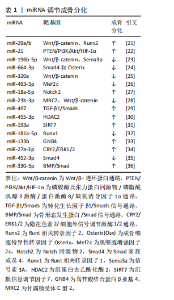
2.1 miRNA与骨质疏松症的研究时间脉络图 骨质疏松症复杂的病理生理过程受到遗传、表观遗传修饰等多种因素的共同调控。其中,表观遗传主要涉及两方面内容:一是基因转录水平选择性表达的调控,如组蛋白修饰、DNA甲基化等;二是基因的转录后调控,包括miRNA、反义RNA、内含子等[12]。在众多的表观遗传机制中,miRNA调控机制是研究较为深入的部分。研究时间脉络图见图3[9,13-17]。 2.2 miRNA作为骨质疏松症治疗靶点的理论基础 miRNA对骨质疏松的调控研究始于2007年[9],至今为止已证明了多条与骨质疏松相关的miRNA,这些miRNA可以通过多方面机制调控骨质疏松的发生,如成骨细胞(骨形成)、破骨细胞(骨吸收)、血管生成等。miRNA作为骨质疏松调控的重要一环,可以作为骨质疏松治疗的重要靶点。此外,研究表明miRNA可以从细胞中分泌,在蛋白质复合物和细胞外囊泡保护下稳定存在于循环系统中[18],因此循环miRNA有可能成为一种骨质疏松的诊断标志物,通过qPCR、基因芯片/测序技术和新兴的等温扩增等技术对体液中miRNA进行检测,在未来可能会成为一种骨质疏松的筛查手段[19]。这里对miRNA从多个角度调控骨质疏松发生的机制进行了总结概述。 2.2.1 miRNA对成骨细胞的调控作用 成骨细胞来源于骨髓间充质干细胞,从骨髓间充质干细胞到成骨细胞的分化程序受多种信号通路[经典Wnt信号通路(Wnt/β-连环蛋白)、骨形态发生蛋白/Smad通路、磷脂酰肌醇3-激酶通路/蛋白激酶B/哺乳动物雷帕霉素靶蛋白通路、丝裂原活化蛋白激酶通路等]和转录因子(Runt相关转录因子2、同源转录因子DLX5、成骨细胞特异性转录因子Osterix、β-连环蛋白、转化生长因子β等)的调节[20]。其中研究最多的信号通路包括Wnt/β-连环蛋白、骨形态发生蛋白/Smad、丝裂原活化蛋白激酶、Hedgehog和Notch信号通路。 大量研究表明,miRNA能够靶向作用于上述信号通路和转录因子,进而实现对成骨细胞分化的调节。例如:miR-29家族miR-29a/b在成骨细胞分化过程中可诱导Runt相关转录因子2和Wnt/β-连环蛋白信号传导来促进成骨分化;miR-29a/c负向调节许多细胞外基质基因表达,抑制成骨分化矿化阶段细胞外基质的组装沉积[21]。miR-21能靶向调控磷酸酶及张力蛋白同源物/磷脂酰肌醇3-激酶通路/蛋白激酶B/缺氧诱导因子1α通路促进骨髓间充质干细胞的迁移和成骨分化[22]。另外,miR-196b-5p可直接靶向信号素3a影响Wnt/β-连环蛋白信号传导,抑制骨形成[23]。研究发现miR-664-3p也可以直接靶Smad4和Osterix抑制成骨和骨形成[24]。更多关于miRNA调控成骨分化的信息见表1[21-36]。"
"

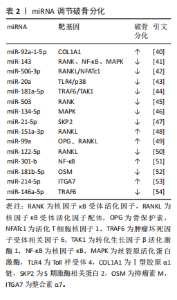
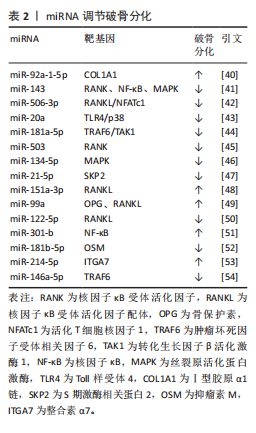
2.2.2 miRNA对破骨细胞的调控作用 破骨细胞源于骨髓中造血干细胞前体,髓系破骨细胞前体在巨噬细胞集落刺激因子和核因子κB受体活化因子配体共同作用下分化为单核破骨细胞,参与破骨细胞生成的主要信号通路有巨噬细胞集落刺激因子/集落刺激因子1受体、核因子κB受体活化因子配体/核因子κB受体活化因子/骨保护素、丝裂原活化蛋白激酶、核因子κB[37]。核因子κB受体活化因子配体/核因子κB受体活化因子/骨保护素信号通路作为骨代谢平衡领域内备受关注的关键调控机制之一,在促进破骨细胞分化与功能活化方面发挥着显著效能[38]。miRNA在调控破骨细胞分化中发挥着重要作用[39]。例如:YU等[40]研究发现miR-92a-1-5p可直接作用于Ⅰ型胶原α1链,增强破骨细胞的分化。HE等[41]发现miR-143通过靶向核因子κB受体活化因子、核因子κB和丝裂原活化蛋白激酶信号通路抑制破骨细胞生成。DINESH等[42]表明,miR-506-3p通过抑制核因子κB受体活化因子配体/活化T细胞核因子1信号通路减少破骨细胞生成。KONG团队[43]研究揭示,miR-20a部分通过Toll样受体4/p38通路抑制破骨细胞的增殖和分化。更多参与破骨细胞分化的miRNA及其靶基因及作用机制总结见表2[40-54]。"

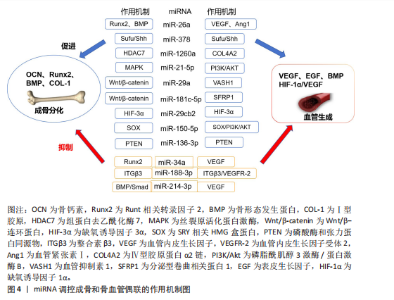
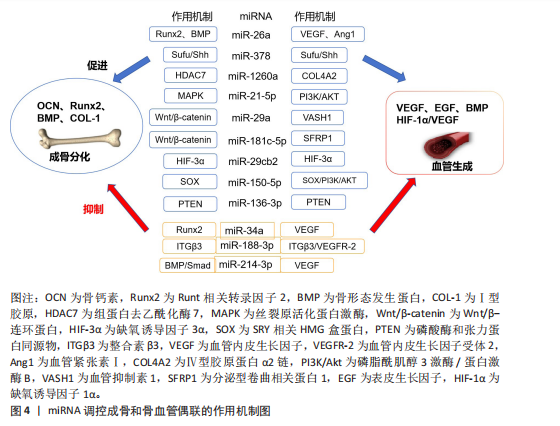
2.2.3 miRNA对骨血管生成的调控 骨是高度血管化的组织,其内部的血管系统不仅输送氧气和必要的营养物质,还参与激素和细胞因子的合成,从而维持骨代谢的稳态[55]。H型血管内皮细胞通过多个信号通路与多种细胞因子的协同作用,刺激骨髓中骨祖细胞的增殖和分化,实现骨形成与血管生成的紧密耦合[56-57]。 骨形成与血管生成的功能偶联受到血管内皮生长因子、骨形态发生蛋白、表皮生长因子样家族成员、缺氧诱导因子1α/血管内皮生长因子、Notch和Wnt/β-连环蛋白信号通路等的调节,研究发现一些miRNA调控血管的生成,参与成骨-血管生成耦合[58]。例如:miR-26a可诱导Runt相关转录因子2和骨形态发生蛋白2刺激成骨分化,也可以靶向骨形态发生蛋白/Smad1信号通路调控血管生成[59]。miR-378可促进骨髓间充质干细胞中的成骨-血管生成偶联,实现潜在的骨再生[60]。骨髓间充质干细胞外泌体中miR-1260a释放可直接靶向组蛋白去乙酰化酶7促进成骨和血管生成[61]。此外,还有许多参与调控血管和骨生成偶联的miRNA,见表3[59-70]。miRNA调控成骨和骨血管偶联的作用机制见图4。"

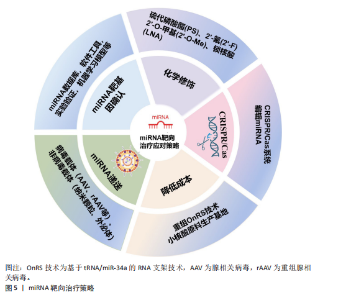
2.3 miRNA靶向治疗骨质疏松症的潜力与挑战 2.3.1 miRNA疗法的应用潜力 miRNA疗法与传统小分子药物相比具有巨大优势,因为约80%疾病相关蛋白靶标是“不可成药”的,而核酸药物突破了蛋白质三维结构限制,miRNA旁分泌效应也提供了更广泛的靶标,使其覆盖整个信号通路[71]。miRNA疗法分为替代疗法(miRNA mimics)和抑制疗法(miRNA inhibitor)[72]。miRNA模拟物可提高胞内miRNA丰度[73]。miRNA抑制剂可特异性靶向内源miRNA,主要方式有miRNA的反义寡核苷酸(AMO),锁定核酸反义寡核苷酸(LNAs)、肽核酸(PNA)、miRNA海绵、miRNA诱饵等[74]。还有基因编辑技术CRISPR/Cas9可在基因水平切割miRNA的Drosha和Dicer加工位点,使miRNA基因组序列发生改变,抑制成熟miRNA合成[75]。 目前已有基于miRNA的多种靶向治疗药物进入临床试验阶段,例如MRX34(miR-34a模拟物)在晚期实体瘤中的Ⅰ期研究已完成[76];Cobomarsen(抗miR-155)针对皮肤T细胞淋巴瘤等肿瘤的Ⅱ期研究正在进行[77];RG-101(miR-122抑制剂)在丙型肝炎治疗中显示出显著病毒载量降低[78];CDR132L(抗miR-132)在心力衰竭治疗中表现出良好的安全性和疗效[79]。上述处于临床前阶段的几种miRNA药物的诞生,证明了miRNA疗法在疾病治疗中有着巨大的应用潜力。 2.3.2 miRNA疗法在骨质疏松症中的应用挑战 近10年关于miRNA与骨质疏松的研究呈快速增长趋势,miRNA作为骨质疏松症的基因治疗靶点展现出显著的潜力,是值得进一步研究的领域[80]。CHEN等[81]研究表明,miR-138-5p通过靶向微管肌动蛋白交联因子1负调控衰老成骨细胞的分化,转染其抑制剂至动物体内可减缓老龄动物的骨量流失。磁性纳米颗粒递送miR-15b-5p至破骨细胞,下调糖尿病骨质疏松大鼠体内神经胶质纤维酸性蛋白表达而抑制破骨细胞分化,减轻骨质疏松[82]。脂肪干细胞来源外泌体含miR-21-5p和let-7b-5p,能够抑制破骨细胞分化,静脉注射脂肪干细胞外泌体可减轻骨质疏松小鼠的骨质流失[83]。此外联合应用miRNA疗法与传统抗骨质疏松药物(如双膦酸盐)可能产生协同效应,为个体化治疗提供新思路,TAIPALEENM?KI等[84]研究表明,miR-19a/b负向调控骨重塑,其拮抗剂既可单独治疗低骨量,又能增强人甲状旁腺激素治疗骨质疏松的疗效。 基于miRNA促进骨再生来治疗骨质疏松症的可行性和有效性已经得到大量细胞和动物研究的支持。但是目前还没有miRNA疗法治疗骨质疏松症的临床研究,主要基于以下几项临床转化挑战,miRNA性质不稳定,易被核酸酶降解,缺乏靶向组织的能力,原料少和制备成本高等问题[85]。 2.4 miRNA靶向治疗骨质疏松症的应对策略 针对miRNA药物在骨质疏松症领域临床转化的挑战,现已开发出一些有效的应对策略,包括精准确定靶基因、化学修饰、利用CRISPR/Cas9编辑miRNA、优化递送系统以及降低生产成本[86]。miRNA的靶向治疗策略见图5。 2.4.1 miRNA靶基因确认 确保miRNA准确靶向其预期的mRNA是miRNA疗法特异性基础。生物信息学预测主要利用miRNA数据库(miRWalk、miRBase、TargetScan等)以及软件工具(MiRcode、Tools4miRs)快速确定miRNA序列和其靶基因;还有多种实验验证方法,包括荧光素酶报告基因实验(Luciferase Assay)、qPCR和Western blot、RNA免疫共沉淀(RIP)、交联免疫沉淀测序(CLIP-seq)等;还有靶向miRNA的CRISPR-Cas9敲除文库(lentiG-miR)有助于系统验证miRNA的作用靶点[87]。随着计算能力指数级的增长和可用数字数据的推动,由数据驱动的大型机器学习模型主导的AI适应不同生物的科学领域,一些用于准确预测人类miRNA靶标的机器学习模型诞生,例如:DeepMirTar(使用卷积神经网络分析miRNA和mRNA的序列特征)[88]。DANE-MDA(通过深度属性网络嵌入预测miRNA与疾病的关联)[89]、miRModuleNet(检测miRNA-mRNA调控模块)[90]。 2.4.2 化学修饰 化学修饰核酸,旨在保护核酸不被降解,增加分子的稳定性。目前,已广泛采用对核酸磷酸"
"
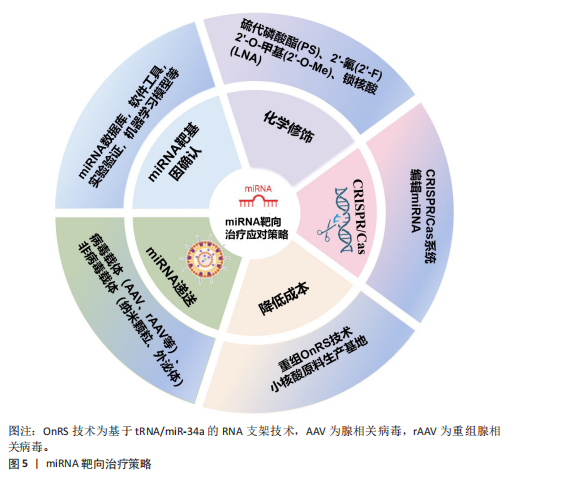
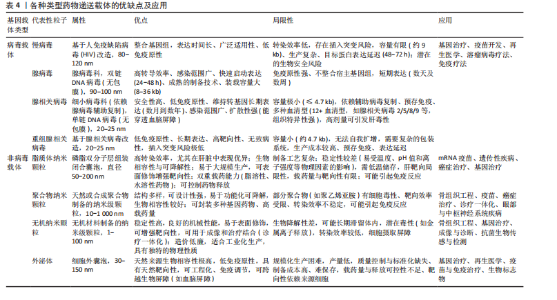
骨架和核糖、碱基的修饰。骨架修饰包括硫代磷酸酯(PS)、肽核酸(PNA)、二酰胺磷酸酯吗啉代(PMO)等;核糖修饰包括2’-O-甲基(2’-O-Me)、2’-氟(2’-F)和锁核酸(LNA)等,常用碱基修饰有2-硫尿苷、5-甲基胞苷等,不论是哪种修饰,均被证实能够有效抵抗核酸酶降解、降低毒性、增强靶点结合力[91]。其中锁核酸(LNA)作为常见的修饰手段,显著增强了对核糖核酸酶的耐药性,并改善了细胞摄取率。若与硫代磷酸酯修饰(PS)组合运用可以获得更好的治疗效果[92]。 2.4.3 CRISPR/Cas9编辑miRNA CRISPR/Cas系统是一种由向导RNA (gRNA)引导的基因编辑平台,可以精准靶向并编辑基因组特定位点。通过CRISPR/Cas技术可以在体内稳定切割miRNA序列以来敲除miRNA,也可以通过调控启动子的方式促进或抑制miRNA的表达[93]。 CRISPR/Cas系统有多种类型,其中最常见的是CRISPR/Cas9。它由Cas9核酸酶与crRNA和tracrRNA制成的嵌合单向导RNA(sgRNA)组成,设计简单、特异性高、操作简便。CRISPR/Cas9可以稳定且特异性地降低miRNA的表达高达96%,其瞬时编辑引起的miRNA敲低表型可在体内外模型中长期稳定维持(长达30 d)[94]。 CRISPR/Cas9系统不仅展现出优越的编辑效能和操作灵活性,还能与高通量测序、单细胞测序等前沿技术相结合,极大地丰富了其应用场景。例如:CRISPR/Cas9与转录组测序技术(RNA-seq)结合,在使用CRISPR/Cas9技术将miRNA敲除或点突变后,采用转录组测序技术分析基因编辑后细胞或模式动物的差异表达基因,同时也可以鉴定出有助于发掘疾病相关的关键突变位点,从而推进疾病的个体化精准治疗研究[95]。 尽管CRISPR/Cas9编辑miRNA具有优势和潜力,但仍面临一些挑战,例如:由于miRNA是短序列,可能出现序列缺失使向导RNA难以检测,向导RNA错配与错误靶标结合而导致的脱靶效应是制约CRISPR/Cas9临床应用的主要障碍之一;此外miRNA的调控机制网络较为复杂,单个miRNA可以调控数百个靶基因,多个miRNA也同时靶向单个基因,因此需要创建多个向导RNA,靶向不同的基因组位点或成熟的miRNA基因组序列才能获得完整的治疗效果;并且,如何将CRISPR/Cas9编辑工具递送到体内目标细胞同时避免或减少脱靶效应仍然是治疗应用的关键障碍。 2.4.4 降低miRNA制作成本 基于miRNA生产成本过高,产业化生产miRNA是目前亟需突破的技术瓶颈。目前RNA生产主要依赖于:①基于DNA的RNA合成;②化学合成的RNA;③体外转录合成的RNA;这些方式价格昂贵、产率低下。加州大学WANG开发了基于tRNA/miR-34a的RNA支架(OnRS)技术在大肠杆菌中合成携带各种小RNA的嵌合RNA的新方法[96],该方法具有高产量、低成本及安全高效的特点[97]。该技术生产的重组miR-129-5p抑制剂能缓解卵巢切除小鼠骨质疏松,效果优于化学合成的抑制剂[98]。另外,国内的瑞博生物公司已投入建成符合GMP标准的小核酸原料公斤级生产基地,能够大幅度降低生产RNA的成本,有利于推动小核酸药物的发展进程。 2.4.5 优化miRNA递送系统 核酸药物的给药方式主要分为全身系统性给药和局部给药(组织特异性递送),miRNA被细胞摄取后,它在细胞质中的生物活性受到多种因素的影响,包括亚细胞定位、与靶标分子的相互作用、细胞微环境等,与药物递送载体偶联是实现miRNA递送的既定方法[99]。常用的递送载体主要分为病毒载体和非病毒载体。 (1)病毒载体:病毒载体主要包括慢病毒(Lentivirus,LV)、腺病毒(Adenovirus,AD)、腺相关病毒(Adeno-associated virus,AAV)及重组腺相关病毒(rAAV)。病毒载体具有优异靶向性和高效转染效率,能有效增加治疗剂在靶细胞浓度,但部分有安全隐患。慢病毒载体包膜或衣壳蛋白易诱导免疫反应限制了疗效。腺病毒免疫原性最强,可致强烈炎症甚至多器官衰竭[100]。腺相关病毒安全性高,宿主基因组整合率极低[101],迄今为止未发现野生型腺相关病毒(wtAAV)致病[102],已获批用于治疗先天性失明(Luxturna)、脊髓性肌萎缩症及血友病B[103]。但是腺相关病毒搭载容量极小(≤4.7 kb),限制了其携带大基因或复杂调控元件的能力。 重组腺相关病毒通过精简腺相关病毒基因组提升安全性,工程改造后可增强组织靶向性,还可通过策略扩大载量[104]。目前OH等[105]使用骨靶向重组腺相关病毒搭载miRNA,降低了Wnt抑制剂schnurri-3和硬化蛋白的表达水平,促进骨生成。在病毒载体方面,重组病毒载体因低免疫原性和高转染效率占据优势,正从实验室走向大规模临床应用,有望应用于骨质疏松症的基因治疗。但重组腺相关病毒无法通过再感染进行自我扩增,生产需要同时表达辅助病毒基因,大规模生产高质量重组腺相关病毒还受到技术限制,通过静脉注射高剂量的重组腺相关病毒会导致肝毒性[104]。 (2)非病毒载体:非病毒载体较病毒载体有多种优势,如原料广泛、化学成分灵活、高载样量可功能化,但也存在转染率低和半衰期短问题,主要有脂质纳米颗粒(lipid nanoparticle,LNP)、聚合物纳米颗粒(Polymeric nanoparticles,PNPs)、无机纳米材料(Inorganic nanomaterials,INP)和外泌体(Exosomes)等[106]。 脂质纳米颗粒:脂质纳米颗粒是由阳离子/可电离脂质、辅助脂质、聚乙二醇脂质、胆固醇4种脂质合成,成分与结构可控,适合规模化生产。不同组合可调节药代动力学与细胞靶向性[107]。脂质纳米颗粒的复杂结构赋予高稳定性,其中阳离子脂质体纳米颗粒可与核酸阴离子结合成稳定复合物,是核酸递送载体中应用最广泛的选择之一[108]。脂质纳米颗粒能控制药物在体内释放的位置和时间,广泛用于疫苗开发,但其靶向性有限,易聚集在肝脏[109]。WANG等[110]发现通过脂质成分优化、靶向配体修饰和微环境响应设计等策略,可制备选择性器官靶向性脂质纳米颗粒以实现组织特异性核酸递送。LIU等[111]发现天冬氨酸(D-Asp8)与羟基磷灰石高度亲和,将天冬氨酸与脂质体偶联,封装miR-148a拮抗剂(抑制破骨细胞生成),可减少骨质疏松小鼠骨吸收。脂质纳米颗粒作为药物递送载体,具有易量产、应用广泛的优势,但存在化学与物理方面的不稳定性,其脂质易水解或封装药物发生泄漏[112]。 聚合物纳米颗粒:聚合物纳米颗粒分为天然和合成两种。天然聚合物包括多糖大分子、蛋白质和透明质酸等,例如:壳聚糖是多糖大分子,因其生物降解性、高生物相容性及功能化特性,有制作可降解骨诱导膜的潜力[113]。合成聚合物为结构可控高分子,分为可生物降解[如聚乙醇酸(PGA)、聚乳酸(PLA)及聚乳酸-羟基乙酸共聚物(PLGA)],获FDA/EMA批准和不可生物降解[如聚乙烯亚胺(PEI)、聚酰胺胺(PAMAM)和聚丙烯亚胺(PPI)]两类[114],可生物降解类在降低免疫原性与细胞毒性方面优势显著。现阶段由聚合物结合无机/有机材料组成的杂化聚合物材料,广泛用于临床试验,包括纳米胶囊、纳米纤维、胶束多种形式,能够结合聚合物和其他分子优势,改变生物分布、溶解度、增强生物相容性[115]。例如:GARCíA-GARCíA等[116]利用脂质-聚合物复合纳米颗粒靶向递送Sfrp-1(Wnt信号抑制剂),促进骨质疏松小鼠骨形成。当前研究聚焦“智能”聚合物纳米颗粒,可响应内/外源性刺激并靶向特定部位,显著提升临床前疗效[117]。实施机器学习可帮助预测聚合物药物的细胞毒性和释放曲线[118]。聚合物纳米颗粒具有高稳定性、低免疫原性及持续递送特性,可功能化设计,广泛应用于生物医学,但制备工艺复杂,在体内代谢清除具有不确定性。 无机纳米颗粒:无机纳米颗粒主要包括金属、碳基、磁性、二氧化硅和磷酸钙等材料,骨骼系统中运用较多的有以下几种,例如:介孔二氧化硅纳米颗粒带正电荷,具有高表面积、有序介孔结构,可控药物释放,表面易功能化修饰,可促进细胞黏附与增殖刺激骨再生[119],对其表面修饰(蛋白涂层和聚合物)可增强稳定性、分散性和生物相容性,对其硅质骨架改造能调节机械强度、降解性能[120];磁性纳米颗粒如Fe3O4和γ-Fe2O3,具有超顺磁性和强磁响应能力,在静磁场作用下可促进骨再生及血管生成[121];磷酸钙纳米颗粒为天然骨矿物质,体内可降解为钙离子参与骨矿化,其优异生物相容性和可降解性使它成为骨科药物递送的理想载体[122];羟基磷灰石钙是骨骼的主要成分,兼具生物相容性、骨传导性及诱导性,带正电荷羟基磷灰石钙纳米颗粒可高效负载核酸,实现骨组织靶向递送[123-124],还可通过掺杂锌、锶、镁等离子进行材料改性,改善成骨和抗菌活性[125]。此外,还有氧化石墨烯[126]、金纳米颗粒[127]、黑鳞纳米片等[128]。无机纳米颗粒通过表面修饰可赋予多功能特性,例如:XIANG等[129]利用抗菌鱼精蛋白修饰磷酸钙纳米颗粒,抗菌的同时可高效递送。仿生纳米技术利用细胞膜包被无机纳米颗粒,具备免疫逃避、长循环及靶向能力[130]。无机纳米颗粒具有亲水性、易修饰、生物相容性和高度稳定性等优点,但不易被机体代谢,有潜在的生物毒性风险[131]。 外泌体:外泌体是直径30-150 nm的细胞外囊泡,由质膜融合释放,参与细胞因子、miRNA等生物分子转运,在细胞通讯中起关键作用,作为天然纳米载体,其低免疫原性、高生物相容性、可穿透多种屏障(血脑屏障),适用于核酸递送[132]。例如:间充质干细胞来源外泌内含多种miRNA和蛋白质,具有天然成骨和血管生成能力,调节免疫反应,倾向于归巢至骨组织,在骨质疏松症治疗中显示出诱人的应用前景[133]。然而天然外泌体的靶向能力通常不够精准,内部成分复杂,治疗效果有限,限制了其临床应用,通过工程化可提高治疗效果,例如:用特定的靶向分子(包括肽)修饰外泌体可实现更精准靶送,通过药物共孵育或基因工程修饰提升载药能力和疗效[134]。例如:WANG等[135]加载骨靶向肽(AspSerSer)6至外泌体构成F6-(DSS)6-exo靶向递送姜黄素至骨组织,恢复骨髓间充质干细胞的成骨分化能力。外泌体凭借生物安全、低免疫原性、可工程化,在药物开发和基因治疗领域表现突出,但面临规模化生产、质控与保存等挑战[136]。随着技术的进展,目前已有人工外泌体结合天然与合成材料,提升载药效率并突破天然细胞外囊泡局限[137],以及人工智能技术优化外泌体分离提纯方法,推动精准递送系统开发,为外泌体治疗开辟新方向,例如:LIM等[138]将人工智能算法与超分辨率显微镜相结合,可显著提高外泌体分析的通量和准确性,提取各种特征参数,并通过统计分析揭示囊泡群体的分布规律和异质性,发现潜在的生物标志物或药物靶点。 各种类型药物递送载体的优缺点及应用,见表4。"

| [1] WU D, CLINE-SMITH A, SHASHKOVA E, et al. T-Cell Mediated Inflammation in Postmenopausal Osteoporosis. Front Immunol. 2021;12:687551. [2] LIANG B, BURLEY G, LIN S, et al. Osteoporosis pathogenesis and treatment: existing and emerging avenues. Cell Mol Biol Lett. 2022;27(1):72. [3] ADEJUYIGBE B, KALLINI J, CHIOU D, et al. Osteoporosis: Molecular Pathology, Diagnostics, and Therapeutics. Int J Mol Sci. 2023;24(19):14583. [4] SINDEL D. Osteoporosis: Spotlight on current approaches to pharmacological treatment. Turk J Phys Med Rehabil. 2023;69(2):140-152. [5] KOMATSU S, KITAI H, SUZUKI HI. Network Regulation of microRNA Biogenesis and Target Interaction. Cells. 2023;12(2):306. [6] ALEXANDRI C, DANIEL A, BRUYLANTS G, et al. The role of microRNAs in ovarian function and the transition toward novel therapeutic strategies in fertility preservation: from bench to future clinical application. Hum Reprod Update. 2020;26(2):174-196. [7] HILL M, TRAN N. miRNA interplay: mechanisms and consequences in cancer. Dis Model Mech. 2021;14(4):dmm047662. [8] NAQVI RA, DATTA M, KHAN SH, et al. Regulatory roles of MicroRNA in shaping T cell function, differentiation and polarization. Semin Cell Dev Biol. 2022; 124:34-47. [9] SUGATANI T, HRUSKA KA. MicroRNA-223 is a key factor in osteoclast differentiation. J Cell Biochem. 2007;101(4):996-999. [10] EMCH MJ, WICIK Z, ASPROS KGM, et al. Estrogen-regulated miRs in bone enhance osteoblast differentiation and matrix mineralization. Mol Ther Nucleic Acids. 2023;33:28-41. [11] SHEN X, ZHU W, ZHANG P, et al. Macrophage miR-149-5p induction is a key driver and therapeutic target for BRONJ. JCI Insight. 2022;7(16):e159865. [12] DEL REAL A, RIANCHO-ZARRABEITIA L, LÓPEZ-DELGADO L, et al. Epigenetics of Skeletal Diseases. Curr Osteoporos Rep. 2018;16(3):246-255. [13] LEE RC, FEINBAUM RL, AMBROS V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75(5):843-854. [14] ESKILDSEN T, TAIPALEENMÄKI H, STENVANG J, et al. MicroRNA-138 regulates osteogenic differentiation of human stromal (mesenchymal) stem cells in vivo. Proc Natl Acad Sci U S A. 2011;108(15):6139-6144. [15] KOCIJAN R, MUSCHITZ C, GEIGER E, et al. Circulating microRNA Signatures in Patients With Idiopathic and Postmenopausal Osteoporosis and Fragility Fractures. J Clin Endocrinol Metab. 2016;101(11):4125-4134. [16] INOUE K, DENG Z, CHEN Y, et al. Bone protection by inhibition of microRNA-182. Nat Commun. 2018;9(1):4108. [17] HWANG JH, PARK YS, KIM HS, et al. Yam-derived exosome-like nanovesicles stimulate osteoblast formation and prevent osteoporosis in mice. J Control Release. 2023;355:184-198. [18] MATSUZAKI J, OCHIYA T. Circulating microRNAs and extracellular vesicles as potential cancer biomarkers: a systematic review. Int J Clin Oncol. 2017;22(3):413-420. [19] TAKIZAWA S, MATSUZAKI J, OCHIYA T. Circulating microRNAs: Challenges with their use as liquid biopsy biomarkers. Cancer Biomark. 2022;35(1):1-9. [20] CAPULLI M, PAONE R, RUCCI N. Osteoblast and osteocyte: games without frontiers. Arch Biochem Biophys. 2014;561:3-12. [21] HORITA M, FARQUHARSON C, STEPHEN LA. The role of miR-29 family in disease. J Cell Biochem. 2021;122(7):696-715. [22] OKA S, LI X, ZHANG F, et al. MicroRNA-21 facilitates osteoblast activity. Biochem Biophys Rep. 2020;25:100894. [23] XIE Y, ZHOU J, TIAN L, et al. miR-196b-5p Regulates Osteoblast and Osteoclast Differentiation and Bone Homeostasis by Targeting SEMA3A. J Bone Miner Res. 2023;38(8):1175-1191. [24] XU Y, JIN Y, HONG F, et al. MiR-664-3p suppresses osteoblast differentiation and impairs bone formation via targeting Smad4 and Osterix. J Cell Mol Med. 2021; 25(11):5025-5037. [25] WANG CG, HU YH, SU SL, et al. LncRNA DANCR and miR-320a suppressed osteogenic differentiation in osteoporosis by directly inhibiting the Wnt/β-catenin signaling pathway. Exp Mol Med. 2020; 52(8):1310-1325. [26] XUE J, LIU L, LIU H, et al. LncRNA SNHG14 activates autophagy via regulating miR-493-5p/Mef2c axis to alleviate osteoporosis progression. Commun Biol. 2023;6(1):1120. [27] HE P, YANG Z, LI H, et al. miR-18a-5p promotes osteogenic differentiation of BMSC by inhibiting Notch2. Bone. 2024; 188:117224. [28] LI R, RUAN Q, YIN F, et al. MiR-23b-3p promotes postmenopausal osteoporosis by targeting MRC2 and regulating the Wnt/β-catenin signaling pathway. J Pharmacol Sci. 2021;145(1):69-78. [29] GU Z, XIE D, HUANG C, et al. MicroRNA-497 elevation or LRG1 knockdown promotes osteoblast proliferation and collagen synthesis in osteoporosis via TGF-β1/Smads signalling pathway. J Cell Mol Med. 2020;24(21):12619-12632. [30] MA H, LI M, JIA Z, et al. MicroRNA-455-3p promotes osteoblast differentiation via targeting HDAC2. Injury. 2022;53(11):3636-3641. [31] SONG CY, GUO Y, CHEN FY, et al. Resveratrol Promotes Osteogenic Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells Through miR-193a/SIRT7 Axis. Calcif Tissue Int. 2022;110(1):117-130. [32] LIU J, CHANG X, DONG D. MicroRNA-181a-5p Curbs Osteogenic Differentiation and Bone Formation Partially Through Impairing Runx1-Dependent Inhibition of AIF-1 Transcription. Endocrinol Metab (Seoul). 2023;38(1):156-173. [33] WANG J, GAO Z, GAO P. MiR-133b Modulates the Osteoblast Differentiation to Prevent Osteoporosis Via Targeting GNB4. Biochem Genet. 2021;59(5):1146-1157. [34] REN LR, YAO RB, WANG SY, et al. MiR-27a-3p promotes the osteogenic differentiation by activating CRY2/ERK1/2 axis. Mol Med. 2021;27(1):43. [35] WU M, WANG H, KONG D, et al. miR-452-3p inhibited osteoblast differentiation by targeting Smad4. PeerJ. 2021;9:e12228. [36] JIN SL, BAI YM, ZHAO BY, et al. Silencing of miR-330-5p stimulates osteogenesis in bone marrow mesenchymal stem cells and inhibits bone loss in osteoporosis by activating Bgn-mediated BMP/Smad pathway. Eur Rev Med Pharmacol Sci. 2020; 24(8):4095-4102. [37] ANWAR A, SAPRA L, GUPTA N, et al. Fine-tuning osteoclastogenesis: An insight into the cellular and molecular regulation of osteoclastogenesis. J Cell Physiol. 2023; 238(7):1431-1464. [38] TOBEIHA M, MOGHADASIAN MH, AMIN N, et al. RANKL/RANK/OPG Pathway: A Mechanism Involved in Exercise-Induced Bone Remodeling. Biomed Res Int. 2020; 2020:6910312. [39] 赵瑞,潘静,裴雪冬,等.破骨细胞相关非编码RNA介导的表观遗传学骨质疏松研究进展[J].中国骨质疏松杂志,2024, 30(4):588-594. [40] YU L, SUI B, FAN W,et al. Exosomes derived from osteogenic tumor activate osteoclast differentiation and concurrently inhibit osteogenesis by transferring COL1A1-targeting miRNA-92a-1-5p.Extracell Vesicles. 2021;10(3):e12056. [41] HE X, ZHU L, AN L, et al. MiR-143 Inhibits Osteoclastogenesis by Targeting RANK and NF-κB and MAPK Signaling Pathways. Curr Mol Pharmacol. 2020;13(3):224-232. [42] DINESH P, KALAISELVAN S, SUJITHA S, et al. miR-506-3p alleviates uncontrolled osteoclastogenesis via repression of RANKL/NFATc1 signaling pathway. J Cell Physiol. 2020;235(12):9497-9509. [43] KONG XH, SHI SF, HU HJ, et al. MicroRNA-20a suppresses RANKL-modulated osteoclastogenesis and prevents bone erosion in mice with rheumatoid arthritis through the TLR4/p38 pathway. J Biol Regul Homeost Agents. 2021;35(3):921-931. [44] XUE HY, LIU MW, YANG G. Resveratrol suppresses lipopolysaccharide-mediated activation of osteoclast precursor RAW 264.7 cells by increasing miR-181a-5p expression. Int J Immunopathol Pharmacol. 2023;37:3946320231154995. [45] HUANG MZ, ZHUANG Y, NING X, et al. Artesunate inhibits osteoclastogenesis through the miR-503/RANK axis. Biosci Rep. 2020;40(7):BSR20194387. [46] HUANG M, WANG Y, WANG Z, et al. miR-134-5p inhibits osteoclastogenesis through a novel miR-134-5p/Itgb1/MAPK pathway. J Biol Chem. 2022;298(7):102116. [47] HUANG Y, YANG Y, WANG J, et al. miR-21-5p targets SKP2 to reduce osteoclastogenesis in a mouse model of osteoporosis. J Biol Chem. 2021;296:100617. [48] HE Y, CHEN D, GUO Q, et al. MicroRNA-151a-3p Functions in the Regulation of Osteoclast Differentiation: Significance to Postmenopausal Osteoporosis. Clin Interv Aging. 2021;16:1357-1366. [49] MOURA SR, BRAS JP, FREITAS J, et al. miR-99a in bone homeostasis: Regulating osteogenic lineage commitment and osteoclast differentiation. Bone. 2020;134:115303. [50] CHOI JH, SUNG SE, KANG KK, et al. Extracellular Vesicles from Human Adipose Tissue-Derived Mesenchymal Stem Cells Suppress RANKL-Induced Osteoclast Differentiation via miR122-5p. Biochem Genet. 2024;62(4):2830-2852. [51] ZHU J, WANG H, LIU H. Osteoclastic miR-301-b knockout reduces ovariectomy (OVX)-induced bone loss by regulating CYDR/NF-κB signaling pathway. Biochem Biophys Res Commun. 2020;529(1):35-42. [52] HAN Z, ZHAN R, CHEN S, et al. miR-181b/Oncostatin m axis inhibits prostate cancer bone metastasis via modulating osteoclast differentiation. J Cell Biochem. 2020;121(2):1664-1674. [53] LIU LL, XIAO YS, HUANG WM, et al. ATF1/miR-214-5p/ITGA7 axis promotes osteoclastogenesis to alter OVX-induced bone absorption. Mol Med. 2022;28(1):56. [54] MINAMI S, FUJII Y, YOSHIOKA Y, et al. Extracellular vesicles from mouse bone marrow macrophages-derived osteoclasts treated with zoledronic acid contain miR-146a-5p and miR-322-3p, which inhibit osteoclast function. Bone. 2025;190:117323. [55] SHANMUGAVADIVU A, BALAGANGADHARAN K, SELVAMURUGAN N. Angiogenic and osteogenic effects of flavonoids in bone regeneration. Biotechnol Bioeng. 2022;119(9):2313-2330. [56] ZHU Y, RUAN Z, LIN Z, et al. The association between CD31hiEmcnhi endothelial cells and bone mineral density in Chinese women. J Bone Miner Metab. 2019;37(6): 987-995. [57] 董晤讯,袁翰,马勇,等.骨血管生成机制与功能的研究进展[J].中国现代医学杂志,2017,27(27):51-58. [58] JIN L, LONG Y, ZHANG Q, et al. MiRNAs regulate cell communication in osteogenesis-angiogenesis coupling during bone regeneration. Mol Biol Rep. 2023;50(10):8715-8728. [59] SADOWSKA JM, ZIMINSKA M, FERREIRA C, et al. Development of miR-26a-activated scaffold to promote healing of critical-sized bone defects through angiogenic and osteogenic mechanisms. Biomaterials. 2023;303:122398. [60] NAN K, ZHANG Y, ZHANG X, et al. Exosomes from miRNA-378-modified adipose-derived stem cells prevent glucocorticoid-induced osteonecrosis of the femoral head by enhancing angiogenesis and osteogenesis via targeting miR-378 negatively regulated suppressor of fused (Sufu). Stem Cell Res Ther. 2021;12(1):331. [61] WU D, CHANG X, TIAN J, et al. Bone mesenchymal stem cells stimulation by magnetic nanoparticles and a static magnetic field: release of exosomal miR-1260a improves osteogenesis and angiogenesis. J Nanobiotechnology. 2021; 19(1):209. [62] QI L, HONG S, ZHAO T, et al. DNA Tetrahedron Delivering miR-21-5p Promotes Senescent Bone Defects Repair through Synergistic Regulation of Osteogenesis and Angiogenesis. Adv Healthc Mater. 2024; 13(30):e2401275. [63] HE WZ, YANG M, JIANG Y, et al. miR-188-3p targets skeletal endothelium coupling of angiogenesis and osteogenesis during ageing. Cell Death Dis. 2022;13(5):494. [64] LU GD, CHENG P, LIU T, et al. BMSC-Derived Exosomal miR-29a Promotes Angiogenesis and Osteogenesis. Front Cell Dev Biol. 2020; 8:608521. [65] ZHA X, SUN B, ZHANG R, et al. Regulatory effect of microRNA-34a on osteogenesis and angiogenesis in glucocorticoid-induced osteonecrosis of the femoral head. J Orthop Res. 2018;36(1):417-424. [66] YU X, RONG PZ, SONG MS, et al. lncRNA SNHG1 induced by SP1 regulates bone remodeling and angiogenesis via sponging miR-181c-5p and modulating SFRP1/Wnt signaling pathway. Mol Med. 2021;27(1):141. [67] OUYANG L, SUN Y, LV D, et al. miR-29cb2 promotes angiogenesis and osteogenesis by inhibiting HIF-3α in bone. iScience. 2021;25(1):103604. [68] WANG X, LI X, LI J, et al. Mechanical loading stimulates bone angiogenesis through enhancing type H vessel formation and downregulating exosomal miR-214-3p from bone marrow-derived mesenchymal stem cells. FASEB J. 2021;35(1):e21150. [69] WU F, SONG C, ZHEN G, et al. Exosomes derived from BMSCs in osteogenic differentiation promote type H blood vessel angiogenesis through miR-150-5p mediated metabolic reprogramming of endothelial cells. Cell Mol Life Sci. 2024;81(1):344. [70] CHEN Y, YU H, ZHU D, et al. miR-136-3p targets PTEN to regulate vascularization and bone formation and ameliorates alcohol-induced osteopenia. FASEB J. 2020;34(4):5348-5362. [71] HUANG CK, KAFERT-KASTING S, THUM T. Preclinical and Clinical Development of Noncoding RNA Therapeutics for Cardiovascular Disease. Circ Res. 2020; 126(5):663-678. [72] LAGGERBAUER B, ENGELHARDT S. MicroRNAs as therapeutic targets in cardiovascular disease. J Clin Invest. 2022; 132(11):e159179. [73] JIN HY, GONZALEZ-MARTIN A, MILETIC AV, et al. Transfection of microRNA Mimics Should Be Used with Caution. Front Genet. 2015;6:340. [74] LIMA JF, CERQUEIRA L, FIGUEIREDO C, et al. Anti-miRNA oligonucleotides: A comprehensive guide for design. RNA Biol. 2018;15(3):338-352. [75] HUSSEN BM, RASUL MF, ABDULLAH SR, et al. Targeting miRNA by CRISPR/Cas in cancer: advantages and challenges. Mil Med Res. 2023;10(1):32. [76] HONG DS, KANG YK, BORAD M, et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br J Cancer. 2020;122(11):1630-1637. [77] SETO AG, BEATTY X, LYNCH JM, et al. Cobomarsen, an oligonucleotide inhibitor of miR-155, co-ordinately regulates multiple survival pathways to reduce cellular proliferation and survival in cutaneous T-cell lymphoma. Br J Haematol. 2018;183(3): 428-444. [78] VAN DER REE MH, DE VREE JM, STELMA F, et al. Safety, tolerability, and antiviral effect of RG-101 in patients with chronic hepatitis C: a phase 1B, double-blind, randomised controlled trial. Lancet. 2017; 389(10070):709-717. [79] TÄUBEL J, HAUKE W, RUMP S, et al. Novel antisense therapy targeting microRNA-132 in patients with heart failure: results of a first-in-human Phase 1b randomized, double-blind, placebo-controlled study. Eur Heart J. 2021;42(2):178-188. [80] YUN L, WANG L, PAN Y, et al. Current status and development trend of miRNAs in osteoporosis-related research: A bibliometric analysis. Folia Histochem Cytobiol. 2021;59(4):203-211. [81] CHEN Z, HUAI Y, CHEN G, et al. MiR-138-5p Targets MACF1 to Aggravate Aging-related Bone Loss. Int J Biol Sci. 2022;18(13):4837-4852. [82] XU C, WANG Z, LIU Y, et al. Delivery of miR-15b-5p via magnetic nanoparticle-enhanced bone marrow mesenchymal stem cell-derived extracellular vesicles mitigates diabetic osteoporosis by targeting GFAP. Cell Biol Toxicol. 2024;40(1):52. [83] LEE KS, LEE J, KIM HK, et al. Extracellular vesicles from adipose tissue-derived stem cells alleviate osteoporosis through osteoprotegerin and miR-21-5p. J Extracell Vesicles. 2021;10(12):e12152. [84] TAIPALEENMÄKI H, SAITO H, SCHRÖDER S, et al. Antagonizing microRNA-19a/b augments PTH anabolic action and restores bone mass in osteoporosis in mice. EMBO Mol Med. 2022;14(11):e13617. [85] DAMMES N, PEER D. Paving the Road for RNA Therapeutics. Trends Pharmacol Sci. 2020;41(10):755-775. [86] BRILLANTE S, VOLPE M, INDRIERI A. Advances in MicroRNA Therapeutics: From Preclinical to Clinical Studies. Hum Gene Ther. 2024;35(17-18):628-648. [87] MERK DJ, PAUL L, TSIAMI F, et al. CRISPR-Cas9 screens reveal common essential miRNAs in human cancer cell lines. Genome Med. 2024;16(1):82. [88] WEN M, CONG P, ZHANG Z, et al. DeepMirTar: a deep-learning approach for predicting human miRNA targets. Bioinformatics. 2018;34(22):3781-3787. [89] JI BY, YOU ZH, WANG Y, et al. DANE-MDA: Predicting microRNA-disease associations via deep attributed network embedding. iScience. 2021;24(6):102455. [90] YOUSEF M, GOY G, BAKIR-GUNGOR B. miRModuleNet: Detecting miRNA-mRNA Regulatory Modules. Front Genet. 2022;13:767455. [91] KHVOROVA A, WATTS JK. The chemical evolution of oligonucleotide therapies of clinical utility. Nat Biotechnol. 2017; 35(3):238-248. [92] WANG J, TAN M, WANG Y, et al. Advances in modification and delivery of nucleic acid drugs. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2023;52(4):417-428. [93] YI B, LARTER K, XI Y. CRISPR/Cas9 System to Knockdown MicroRNA In Vitro and In Vivo. Methods Mol Biol. 2021;2300:133-139. [94] CHUNG PJ, CHUNG H, OH N, et al. Efficiency of Recombinant CRISPR/rCas9-Mediated miRNA Gene Editing in Rice. Int J Mol Sci. 2020;21(24):9606. [95] SELVAKUMAR SC, PREETHI KA, ROSS K, et al. CRISPR/Cas9 and next generation sequencing in the personalized treatment of Cancer. Mol Cancer. 2022;21(1):83. [96] WANG WP, HO PY, CHEN QX, et al. Bioengineering Novel Chimeric microRNA-34a for Prodrug Cancer Therapy: High-Yield Expression and Purification, and Structural and Functional Characterization. J Pharmacol Exp Ther. 2015;354(2):131-141. [97] CHEN QX, WANG WP, ZENG S, et al. A general approach to high-yield biosynthesis of chimeric RNAs bearing various types of functional small RNAs for broad applications. Nucleic Acids Res. 2015;43(7):3857-3869. [98] YIN C, TIAN Y, YU Y, et al. miR-129-5p Inhibits Bone Formation Through TCF4. Front Cell Dev Biol. 2020;8:600641. [99] RUSESKA I, ZIMMER A. Cellular uptake and trafficking of peptide-based drug delivery systems for miRNA. Eur J Pharm Biopharm. 2023;191:189-204. [100] WANG X, MA C, RODRÍGUEZ LABRADA R, et al. Recent advances in lentiviral vectors for gene therapy. Sci China Life Sci. 2021;64(11):1842-1857. [101] AL-HEETI OM, CATHRO HP, ISON MG. Adenovirus Infection and Transplantation. Transplantation. 2022;106(5):920-927. [102] COSTA VERDERA H, KURANDA K, MINGOZZI F. AAV Vector Immunogenicity in Humans: A Long Journey to Successful Gene Transfer. Mol Ther. 2020;28(3):723-746. [103] MENDELL JR, AL-ZAIDY SA, RODINO-KLAPAC LR, et al. Current Clinical Applications of In Vivo Gene Therapy with AAVs. Mol Ther. 2021;29(2):464-488. [104] WANG JH, GESSLER DJ, ZHAN W, et al. Adeno-associated virus as a delivery vector for gene therapy of human diseases. Signal Transduct Target Ther. 2024;9(1):78. [105] OH WT, YANG YS, XIE J, et al. WNT-modulating gene silencers as a gene therapy for osteoporosis, bone fracture, and critical-sized bone defects. Mol Ther. 2023;31(2):435-453. [106] WANG C, PAN C, YONG H, et al. Emerging non-viral vectors for gene delivery. J Nanobiotechnology. 2023;21(1):272. [107] PAUNOVSKA K, LOUGHREY D, DAHLMAN JE. Drug delivery systems for RNA therapeutics. Nat Rev Genet. 2022;23(5):265-280. [108] KOYNOVA R, TENCHOV B. Cationic lipids: molecular structure/ transfection activity relationships and interactions with biomembranes. Top Curr Chem. 2010; 296:51-93. [109] SCHOENMAKER L, WITZIGMANN D, KULKARNI JA, et al. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int J Pharm. 2021;601:120586. [110] WANG X, LIU S, SUN Y, et al. Preparation of selective organ-targeting (SORT) lipid nanoparticles (LNPs) using multiple technical methods for tissue-specific mRNA delivery. Nat Protoc. 2023;18(1):265-291. [111] LIU J, DANG L, LI D, et al. A delivery system specifically approaching bone resorption surfaces to facilitate therapeutic modulation of microRNAs in osteoclasts. Biomaterials. 2015;52:148-160. [112] FAN Y, MARIOLI M, ZHANG K. Analytical characterization of liposomes and other lipid nanoparticles for drug delivery. J Pharm Biomed Anal. 2021;192:113642. [113] FARJAMINEJAD S, FARJAMINEJAD R, GARCIA-GODOY F. Nanoparticles in Bone Regeneration: A Narrative Review of Current Advances and Future Directions in Tissue Engineering. J Funct Biomater. 2024;15(9):241. [114] CHENTHAMARA D, SUBRAMANIAM S, RAMAKRISHNAN SG, et al. Therapeutic efficacy of nanoparticles and routes of administration. Biomater Res. 2019;23:20. [115] FERREIRA SOARES DC, DOMINGUES SC, VIANA DB, et al. Polymer-hybrid nanoparticles: Current advances in biomedical applications. Biomed Pharmacother. 2020;131:110695. [116] GARCÍA-GARCÍA P, REYES R, GARCÍA-SÁNCHEZ D, et al. Nanoparticle-mediated selective Sfrp-1 silencing enhances bone density in osteoporotic mice. J Nanobiotechnology. 2022;20(1):462. [117] BEACH MA, NAYANATHARA U, GAO Y, et al. Polymeric Nanoparticles for Drug Delivery. Chem Rev. 2024;124(9):5505-5616. [118] GREENBERG ZF, GRAIM KS, HE M. Towards artificial intelligence-enabled extracellular vesicle precision drug delivery. Adv Drug Deliv Rev. 2023;199:114974. [119] PINNA A, TORKI BAGHBADERANI M, VIGIL HERNÁNDEZ V, et al. Nanoceria provides antioxidant and osteogenic properties to mesoporous silica nanoparticles for osteoporosis treatment. Acta Biomater. 2021;122:365-376. [120] KANKALA RK, HAN YH, NA J, et al. Nanoarchitectured Structure and Surface Biofunctionality of Mesoporous Silica Nanoparticles. Adv Mater. 2020; 32(23):e1907035. [121] XIA Y, SUN J, ZHAO L, et al. Magnetic field and nano-scaffolds with stem cells to enhance bone regeneration. Biomaterials. 2018;183:151-170. [122] KHALIFEHZADEH R, ARAMI H. Biodegradable calcium phosphate nanoparticles for cancer therapy. Adv Colloid Interface Sci. 2020;279:102157. [123] KAWSAR M, SAHADAT HOSSAIN M, ALAM MK, et al. Synthesis of pure and doped nano-calcium phosphates using different conventional methods for biomedical applications: a review. J Mater Chem B. 2024;12(14):3376-3391. [124] MUNIR MU, SALMAN S, JAVED I, et al. Nano-hydroxyapatite as a delivery system: overview and advancements. Artif Cells Nanomed Biotechnol. 2021;49(1):717-727. [125] SHOKRI M, KHARAZIHA M, TAFTI HA, et al. Synergic role of zinc and gallium doping in hydroxyapatite nanoparticles to improve osteogenesis and antibacterial activity. Biomater Adv. 2022;134:112684. [126] ITOO AM, VEMULA SL, GUPTA MT, et al. Multifunctional graphene oxide nanoparticles for drug delivery in cancer. J Control Release. 2022;350:26-59. [127] GUPTA A, SINGH S. Multimodal Potentials of Gold Nanoparticles for Bone Tissue Engineering and Regenerative Medicine: Avenues and Prospects. Small. 2022;18(29): e2201462. [128] XU Y, CHEN S, ZHANG Y, et al. Antibacterial black phosphorus nanosheets for biomedical applications. J Mater Chem B. 2023;11(30):7069-7093. [129] XIANG C, TENKUMO T, OGAWA T, et al. Gene transfection achieved by utilizing antibacterial calcium phosphate nanoparticles for enhanced regenerative therapy. Acta Biomater. 2021;119:375-389. [130] SONG W, JIA P, ZHANG T, et al. Cell membrane-camouflaged inorganic nanoparticles for cancer therapy. J Nanobiotechnology. 2022;20(1):289. [131] 秦灿,梁爱玲,刘勇军.纳米载体在抗癌肽递送中的应用[J].中国生物化学与分子生物学报,2025,41(6):833-842. [132] ESCUDÉ MARTINEZ DE CASTILLA P, TONG L, HUANG C, et al. Extracellular vesicles as a drug delivery system: A systematic review of preclinical studies. Adv Drug Deliv Rev. 2021;175:113801. [133] LI X, SI Y, LIANG J, et al. Enhancing bone regeneration and immunomodulation via gelatin methacryloyl hydrogel-encapsulated exosomes from osteogenic pre-differentiated mesenchymal stem cells. J Colloid Interface Sci. 2024;672:179-199. [134] ZHANG M, HU S, LIU L, et al. Engineered exosomes from different sources for cancer-targeted therapy. Signal Transduct Target Ther. 2023;8(1):124. [135] WANG Y, SUN L, DONG Z, et al. Targeted inhibition of ferroptosis in bone marrow mesenchymal stem cells by engineered exosomes alleviates bone loss in smoking-related osteoporosis. Mater Today Bio. 2025;31:101501. [136] ISAAC R, REIS FCG, YING W, et al. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab. 2021;33(9):1744-1762. [137] LI YJ, WU JY, LIU J, et al. Artificial exosomes for translational nanomedicine. J Nanobiotechnology. 2021;19(1):242. [138] LIM HJ, KIM GW, HEO GH, et al. Nanoscale single-vesicle analysis: High-throughput approaches through AI-enhanced super-resolution image analysis. Biosens Bioelectron. 2024;263:116629. |
| [1] | Zhang Haiwen, Zhang Xian, Xu Taichuan, Li Chao. Bibliometric and visual analysis of the research status and trends of senescence in osteoporosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(6): 1580-1591. |
| [2] | Wang Yan, Lyu Hao, Hu Zhimu, Zhou Yao, Liu Qiang, Yang Yuxiang, Yi Hairu, Wang Jiuxiang, Jiang Ting. Intervention with Compound Kidney-Invigorating Granules in a mouse model of osteoporosis: role of the TRIB3/beta-catenin axis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(23): 6142-6149. |
| [3] | Du Xingbin, Jiang Fugao, Kong Jianda. Traditional Chinese sports in the treatment of osteoporosis: potential biological mechanisms and clinical application progress [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(23): 5943-5953. |
| [4] | Gao Jiabin, Li Tianqi, Xu Kun, Zhu Hanmin, Zhou Xi, Li Wei. Mitophagy regulates osteoclasts: a new perspective for osteoporosis treatment [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(23): 5982-5991. |
| [5] | Wang Xinyue, Li Hongli, Guo Chunhui, Chen Jibing, Yu Hua. Changes in the expression of six microRNAs in ovarian tissue from animal models of premature ovarian failure and in peripheral blood of patients with premature ovarian failure [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(18): 4675-4684. |
| [6] | Wang Siwei, Yao Xiaosheng, Qi Xiaonan, Wang Yu, Cui Haijian, Zhao Jiaxuan. Matrix metalloproteinase 9 mediates mitophagy to regulate osteogenesis and myogenesis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(18): 4557-4567. |
| [7] | Zhao Yu, Xue Yun, Huang Jiajun, Wu Diyou, Yang Bin, Huang Junqing. Total flavonoids from Semen Cuscutae inhibits osteoblast apoptosis in hormone-induced femoral head avascular necrosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(17): 4289-4298. |
| [8] | Fu Jingyue, Zhou Qinfeng, Li Muzhe, Ma Yong, Pan Yalan, Sun Jie, Huang Xiangyang, Guo Yang. Preparation and evaluation of an animal model of osteoporosis and osteoarthritis comorbidity in rats [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(17): 4299-4308. |
| [9] | Sun Long, Wu Haiyang, Tong Linjian, Liu Rui, Yang Weiguang, Xiao Jian, Liu Lice, Sun Zhiming. Regulatory mechanism of leptin in bone metabolism [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(12): 3100-3108. |
| [10] | Shi Tengbo, Tang Yanfeng, Zhang Mengyu, Wang Xingfei, Li Chenyang, Shi Jinyu, Guo Chaowei, Li Yanzhou, He Zike, Wang Shangzeng. Effect of high-dose low molecular weight heparin on the healing of femoral shaft fractures [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(12): 2957-2964. |
| [11] | Lu Xiuli, Xu Huazhen, Chen Yuxing, Yao Nan, Hu Zixuan, Huang Dane. Mechanism of Jiangu Formula in treating osteoporosis based on osteoclast-osteoblast coupling [J]. Chinese Journal of Tissue Engineering Research, 2025, 29(32): 6828-6835. |
| [12] | Wu Wangxiang, Ran Dongcheng, Xu Jiamu, Xu Jiafu, Chen Jingjing, Wang Chunqing. Long noncoding RNAs related to osteoporosis: current research status and developmental trends [J]. Chinese Journal of Tissue Engineering Research, 2025, 29(29): 6360-6368. |
| [13] | Tang Zhi, Shao Yang, Li Shaoshuo, Qi Shubin, Lu Hengyang, Wu Mao, Yang Junfeng, Wang Jianwei. Single-cell sequencing reveals heterogeneity of B cells in osteoporosis patients and their interactions with osteoblasts [J]. Chinese Journal of Tissue Engineering Research, 2025, 29(26): 5501-5510. |
| [14] | Wang Lei, Wang Baiyan, Zhou Chunguang, Ren Xiaoyun, Dai Yueyou, Feng Shuying. Role of different cell-derived exosomal miRNAs in progression, diagnosis, and prognosis of gastric cancer [J]. Chinese Journal of Tissue Engineering Research, 2025, 29(25): 5434-5442. |
| [15] | Li Haishan, Wu Yuheng, Liang Zixuan, Zhang Shiyin, Zhang Zhen, Mai Bin, Deng Wei, Li Yongxian, Tang Yongchao, , Zhang Shuncong, , Yuan Kai, . Carnosic acid inhibits osteoclast differentiation by inhibiting mitochondrial activity [J]. Chinese Journal of Tissue Engineering Research, 2025, 29(2): 245-253. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||