Chinese Journal of Tissue Engineering Research ›› 2026, Vol. 30 ›› Issue (35): 9239-9247.doi: 10.12307/2026.468
Previous Articles Next Articles
Type VI collagen: a multifunctional regulator in bone homeostasis and tissue engineering
Dong Shiming1, 2, Xie Zhenzi3, Tao Rongrong4, Mardan·Mamat2, Ma Hairong1, 5
- 1Research Institute of Clinical Medicine, 2Department of Spine Surgery, The First Affiliated Hospital of Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China; 3School of Basic Medical Sciences, Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China; 4Infectious Diseases Department, Binhu Hospital, Hefei First People’s Hospital, Hefei 230092, Anhui Province, China; 5School of Pharmacy, Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China
-
Received:2025-11-18Revised:2026-03-02Online:2026-12-18Published:2026-04-28 -
Contact:Ma Hairong, MD, Researcher, Research Institute of Clinical Medicine, The First Affiliated Hospital of Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China; School of Pharmacy, Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China Co-corresponding author: Mardan·Mamat, MS, Chief physician, Department of Spine Surgery, The First Affiliated Hospital of Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China -
About author:Dong Shiming, MS candidate, Research Institute of Clinical Medicine, The First Affiliated Hospital of Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China; Department of Spine Surgery, The First Affiliated Hospital of Xinjiang Medical University, Urumqi 830054, Xinjiang Uygur Autonomous Region, China -
Supported by:National Natural Science Foundation of China, No. 82060411 (to MHR); Key Project of the Xinjiang Uygur Autonomous Region Natural Science Foundation, No. 2021D01D21 (to MHR)
CLC Number:
Cite this article
Dong Shiming, Xie Zhenzi, Tao Rongrong, Mardan·Mamat, Ma Hairong. Type VI collagen: a multifunctional regulator in bone homeostasis and tissue engineering[J]. Chinese Journal of Tissue Engineering Research, 2026, 30(35): 9239-9247.
share this article
Add to citation manager EndNote|Reference Manager|ProCite|BibTeX|RefWorks
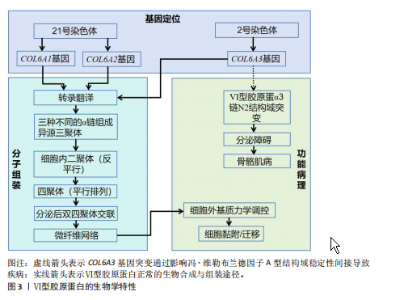
2.1 Ⅵ型胶原蛋白的结构、功能及生物学特性 2.1.1 Ⅵ型胶原蛋白的染色体分子结构和定位 Ⅵ型胶原蛋白是一种结构独特的细胞外基质成分,典型分子结构为由α1、α2和α3链构成的异源三聚体,该三聚体包含1个较短的三螺旋结构域和2个较大的球状结构域,这些球状结构域的结构与功能不仅受可变剪接的调控,还受到细胞内外的广泛翻译后修饰的影响。Ⅵ型胶原蛋白的组装始于细胞内:首先异源三聚体单体组装成反平行的二聚体,随后二聚体以平行方式排列形成四聚体,这种独特的组装模式是维持结构稳定性和复杂生物学功能的结构基础。Ⅵ型胶原蛋白凭借特殊的分子构象,在调控细胞黏附、迁移与增殖等过程中发挥重要作用,尤其在骨骼与软骨组织中扮演着关键角色,因此成为骨骼系统研究的重要靶点[2,5]。 此外,Ⅵ型胶原蛋白的分子结构具备一定的适应性,能够在不同生理和病理条件下发生构象变化,从而动态调节生物学功能及其与其他细胞外基质分子的相互作用[6]。通过整合X射线晶体学、细胞实验和小角X射线散射技术,解析Ⅵ型胶原蛋白α3链N2 VWA结构域(von Willebrand因子A结构域)的三维结构;细胞实验显示,该结构域的突变可显著降低Ⅵ型胶原蛋白分泌效率;小角X射线散射分析进一步证实该结构域在溶液中呈串联模块化排列,并表现出动态构象变化[7]。该研究提示,Ⅵ型胶原蛋白α3链N2 VWA结构域固有的构象可塑性,可能通过多价结合的方式,协同调节Ⅵ型胶原蛋白与细胞外基质的相互作用及其介导的力学信号传导。 对从人类胎盘中分离并酶解的Ⅵ型胶原蛋白进行分析,明确了其中各α链的基因染色体定位:α1(Ⅵ)和α2(Ⅵ)链的基因定位于人类21号染色体,而α3(Ⅵ)链的基因则定位于2号染色体[8-9]。这些发现为理解Ⅵ型胶原蛋白在细胞间的相互作用和组织修复等生物学过程中的功能提供了重要线索。见图3。 2.1.2 Ⅵ型胶原蛋白的生物合成与降解 Ⅵ型胶原蛋白的合成始于3条α链,它们分别由COL6A1、COL6A2和COL6A3基因编码,这些α链在粗面内质网中翻译后,经历脯氨酸、赖氨酸的羟基化以及羟赖氨酸的糖基化等翻译后修"
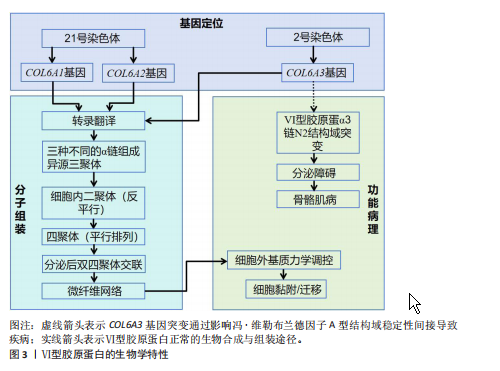
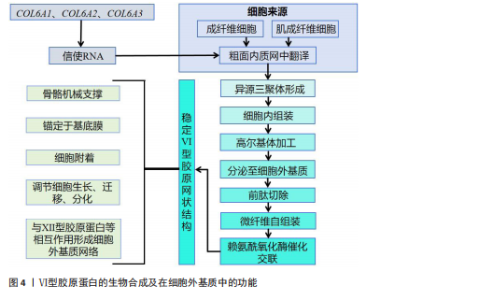
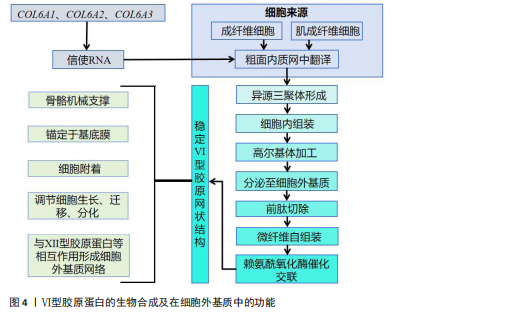
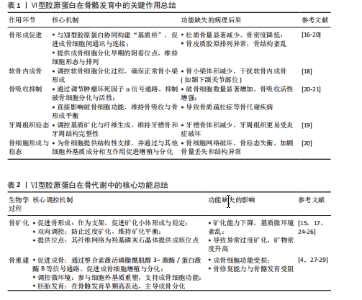
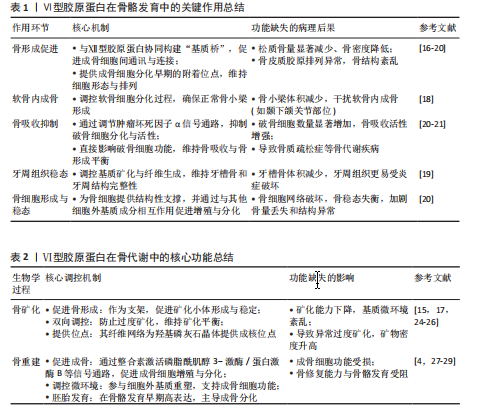
饰,最终组装成异源三聚体三螺旋结构,随后这些三聚体在细胞内组装成反平行二聚体,并进一步聚合为四聚体,最后通过高尔基体被分泌到细胞外,前胶原分子两端的球状前肽被蛋白酶切除,成熟的Ⅵ型胶原蛋白通过非共价相互作用自组装为微纤维,并在赖氨酰氧化酶催化下形成分子间共价交联,最终构建稳定的网状结构,锚定于基底膜,参与骨骼的机械支撑,该过程主要由成纤维细胞和肌成纤维细胞介导,羟基化不足或交联缺陷可导致胶原结构异常[10-11]。在降解过程中,Ⅵ型胶原蛋白主要被基质金属蛋白酶降解;基质金属蛋白酶可特异性切割Ⅵ型胶原蛋白,从而调控它在细胞外基质中的稳定性与功能。Ⅵ型胶原蛋白的异常降解与多种疾病密切相关,尤其是乌尔里希型先天性肌营养不良、其他类型的肌营养不良及骨关节炎等。以乌尔里希先天性肌营养不良为例,COL6A1、COL6A2或COL6A3基因突变,尤其是显性杂合突变,可通过显性负效应破坏Ⅵ型胶原蛋白的正常组装,并加剧胶原蛋白降解,导致细胞外基质稳态失衡,进而引发特征性病理改变[12]。综上所述,Ⅵ型胶原蛋白通过合成(依赖于α链的正确组装与分子交联)与降解(基质金属蛋白酶介导)之间的精密调控,维持骨骼基质的稳态与机械支撑功能,核心病理机制在于显性杂合突变通过显性负效应破坏Ⅵ型胶原蛋白的正常组装并加剧其降解,导致细胞外基质结构破坏,从而引发乌尔里希先天性肌营养不良等疾病。因此,深入解析Ⅵ型胶原蛋白合成与降解的调控网络,将为相关疾病的靶向治疗提供关键靶点。 2.1.3 Ⅵ型胶原蛋白在细胞外基质中的功能 Ⅵ型胶原蛋白在细胞外基质中具有多重生物学功能,尤其在维持组织结构完整性与功能稳态方面发挥关键作用。它不仅为细胞附着提供结构基础,还参与细胞信号转导,调控细胞的增殖、迁移与分化。ANTONIEL等[12]通过对2例患者的肌腱活检样本及体外培养细胞进行超微结构观察、生物化学分析和RT-qPCR检测,揭示了Ⅵ型胶原蛋白缺陷与肌肉功能障碍之间的关联。研究表明,Ⅵ型胶原蛋白可与多种基质成分(如大分子蛋白聚糖和糖胺聚糖)协同作用,构建复杂的细胞外基质网络,进而调控细胞的机械信号转导及生物学响应[13]。在肌肉和肌腱等组织中,Ⅵ型胶原蛋白的缺失或突变可导致细胞极性丧失及迁移能力显著降低,进而损害组织的正常功能与修复潜能[14]。近期研究表明,细丝蛋白B(Filamin B,FLNB)通过多维度调控机制影响Ⅵ型胶原蛋白的表达与功能,为阐明骨骼疾病的发生机制提供了新视角。首先,细丝蛋白B敲除可显著下调COL6A2与COL6A3的mRNA表达水平,该结果经RNA测序和RT-qPCR验证,提示细丝蛋白B可能通过直接或间接途径调控Ⅵ型胶原蛋白的基因转录;进一步分析表明,细丝蛋白B可能通过磷脂酰肌醇3-激酶/蛋白激酶B(phosphoinositide 3-kinase/protein kinase B,PI3K/AKT)信号通路间接影响Ⅵ型胶原蛋白的合成,或作为RNA结合蛋白直接作用于Ⅵ型胶原蛋白的mRNA稳定性及剪接过程,从而调节Ⅵ型胶原蛋白表达水平[15]。此外,Ⅵ型胶原蛋白还可与Ⅻ型胶原蛋白等其他胶原亚型相互作用,形成功能性复合物,进一步强化Ⅵ型胶原蛋白在组织结构维持与修复中的作用[6]。综上所述,Ⅵ型胶原蛋白通过维持骨骼系统细胞外基质的力学稳定性并调控细胞分化,在骨发育过程中发挥关键作用。细丝蛋白B可调控Ⅵ型胶原蛋白表达的发现,为骨相关疾病的治疗提供了新的潜在靶点。未来的研究需致力于全面解析Ⅵ型胶原蛋白的动态表达规律,积极探索靶向细丝蛋白B-Ⅵ型胶原蛋白信号轴的骨组织再生策略,并推动基于Ⅵ型胶原蛋白的生物材料临床应用,以期提升骨组织修复的疗效。见图4。 2.2 Ⅵ型胶原蛋白在骨骼发育中的作用 2.2.1 Ⅵ型胶原蛋白在骨形成中的促进作用 Ⅵ型胶原蛋白是骨组织细胞外基质中不可或缺的成分,在骨形成过程中发挥关键的调控作用。具体而言,Ⅵ型胶原蛋白通过精确调控成骨细胞行为、促进细胞间通讯以及维持骨基质结构的完整性,驱动正常的骨生成过程,并有助于防止骨量丢失和骨脆性增加,作用机制主要包括:其一,Ⅵ型胶原蛋白可与Ⅻ型胶原蛋白协同作用,在成骨细胞之间形成“基质桥”,从而显著促进细胞连接与信号交流,为骨形成所必需的细胞网络构建奠定基础[16];其二,成骨细胞在分化早期高度依赖Ⅵ型胶原蛋白所提供"
的附着位点;若Ⅵ型胶原蛋白缺失,则会导致成骨细胞形态异常与排列紊乱,进而破坏骨基质的正常结构。研究表明,Ⅵ型胶原蛋白缺失会对骨形成产生多方面的负面影响,主要体现在以下几个方面:①骨量与骨密度:在Ⅵ型胶原蛋白缺陷的小鼠模型中,可观察到松质骨量显著减少、骨密度降低,并伴有骨皮质胶原纤维排列异常,这凸显了Ⅵ型胶原蛋白对于维持骨量及骨结构完整性的关键作用[17-20];②软骨内成骨:在颞下颌关节等特定部位,Ⅵ型胶原蛋白缺失引发软骨细胞分化障碍,导致骨小梁体积减少,干扰软骨内成骨过程[18];③骨吸收调节:Ⅵ型胶原蛋白缺失通过增强肿瘤坏死因子α信号通路,显著促进破骨细胞分化及骨吸收活性;④牙槽骨与牙周组织:Ⅵ型胶原蛋白调控基质矿化与纤维生成,其缺失会导致牙槽骨体积减少,并使牙周组织更易受炎症破坏[19]。值得注意的是,Ⅵ型胶原蛋白的功能在不同骨部位可能存在差异,这与所处环境的特定损伤状态和生物力学微环境相关。特别值得注意的是,Ⅵ型胶原蛋白在骨细胞形成过程中扮演核心角色,不仅为骨细胞提供结构性支撑,还通过与其他细胞外基质成分的相互作用,促进骨细胞的增殖与分化[20]。现有证据表明,Ⅵ型胶原蛋白通过促进成骨细胞的有效通讯、调控骨基质结构以及抑制过度骨吸收,共同维持骨组织的正常形成与稳态平衡;反之,Ⅵ型胶原蛋白功能缺失则会直接导致骨量减少、骨结构紊乱及骨吸收活动增强。 综上所述,Ⅵ型胶原蛋白在骨形成中并非仅作为静态的结构组分,而是一个动态调控骨发育与稳态的核心分子枢纽,作用机制涉及细胞通讯网络的构建、成骨细胞功能的精密调控、骨吸收平衡的维持以及骨基质矿化与结构稳定性的支撑。 2.2.2 Ⅵ型胶原蛋白对骨吸收的抑制作用 Ⅵ型胶原蛋白通过与破骨细胞等骨代谢相关细胞相互作用,参与骨代谢的调控。基于Col6a2基因敲除小鼠的转录组分析,Ⅵ型胶原蛋白缺失可通过增强破骨细胞的骨吸收活性,导致小鼠发生骨质疏松,该表型与骨骼中破骨细胞相关基因表达上调相一致[20]。进一步研究发现,在骨髓间充质干细胞中,敲除Col6a2基因后,在肿瘤坏死因子α刺激下,细胞中破骨细胞生成相关mRNA的表达水平显著高于野生型细胞;同时,肿瘤坏死因子α下游信号靶标磷酸化p65水平也明显升高。这些结果提示,Col6a2基因缺失可增强骨髓间充质干细胞对肿瘤坏死因子α信号的敏感性,从而促进破骨细胞的生成。此外,Ⅵ型胶原蛋白还可直接影响破骨细胞的分化与功能活性,参与调控骨吸收与骨形成之间的动态平衡。在Ⅵ型胶原蛋白缺失的小鼠模型中,破骨细胞数量显著增加,表明Ⅵ型胶原蛋白在抑制破骨细胞活性中发挥关键作用[21]。破骨细胞的异常活化与数量增多是骨质疏松症等骨代谢性疾病的关键病理特征。因此,系统解析Ⅵ型胶原蛋白调控破骨细胞分化与功能的分子机制,对于阐明骨骼稳态失衡的分子基础、开发靶向治疗策略具有重要的科学意义。见表1。 2.2.3 Ⅵ型胶原蛋白参与骨矿化的生物学过程 骨矿化是一个受多阶段精密调控的生物学过程,钙离子与磷酸根离子经基质囊泡运输并沉积形成矿化结节,其中矿化小体作为核心功能单位,其形成与稳定性直接决定骨基质的矿化效率[21-23]。在此过程中,Ⅵ型胶原蛋白通过与基质成分(如镁掺杂羟基磷灰石)相互作用,直接促进矿化小体的形成并增强结构稳定性,从而推动矿化进程[24]。WANG等[15]研究表明,Ⅵ型胶原蛋白表达下调可显著削弱成骨细胞的黏附、增殖与分化能力,导致细胞外基质微环境紊乱,进而抑制矿化潜能。值得注意的是,Ⅵ型胶原蛋白在骨矿化过程中亦具有负向调控作用:研究表明,Ⅵ型胶原蛋白可通过调节细胞外基质的组织特性,抑制牙槽骨与牙本质的过度矿化;相反,在Ⅵ型胶原蛋白缺失的小鼠模型中,矿物密度显著升高,提示Ⅵ型胶原蛋白在生物矿化中发挥负调控功能[17]。进一步研究发现,Ⅵ型胶原蛋白纤维间的间隙可为羟基磷灰石晶体提供成核位点,并通过与特定成核抑制因子协同作用,动态调控矿化进程[25]。此外,Ⅵ型胶原蛋白自身的磷酸化修饰可引入新的成核位点,促进羟基磷灰石在胶原纤维内的有序分层沉积,从而优化矿化结构的构建[26]。在这一精细调控过程中,非胶原蛋白通过其富含阴离子的结构域介导矿物质沉积,并与胶原蛋白协同促进纤维矿化。 综上所述,Ⅵ型胶原蛋白通过调控细胞外基质微环境、介导羟基磷灰石晶体成核以及协同矿化功能单元的组装等多重机制,在骨矿化过程中发挥不可或缺的作用。 2.2.4 Ⅵ型胶原蛋白促进骨重建的分子机制 Ⅵ型胶原蛋白通过多种分子机制调控成骨细胞的增殖与分化。首先,Ⅵ型胶原蛋白可与细胞表面整合素结合,激活下游信号通路,进而促进成骨细胞增殖。例如,Ⅵ型胶原蛋白与整合素结合可激活磷脂酰肌醇3-激酶/蛋白激酶B信号通路,该通路在调控细胞增殖与存活中发挥关键作用[27];其次,Ⅵ型胶原蛋白可通过调节生长因子的分泌,进一步促进成骨细胞分化。研究表明,Ⅵ型胶原蛋白可促进成骨细胞分泌成纤维细胞生长因子,后者进一步刺激成骨细胞的增殖与分化[28]。此外,Ⅵ型胶原蛋白还可通过与细胞表面受体神经-胶质抗原2相互作用,调控成骨细胞分化[29]。同时,Ⅵ型胶原蛋白参与细胞外基质重塑,为成骨细胞提供适宜的微环境,从而支持成骨细胞功能发挥。上述机制表明,Ⅵ型胶原蛋白在骨骼修复过程中不仅促进成骨细胞增殖,亦增强成骨细胞分化能力;反之,Ⅵ型胶原蛋白缺失可导致成骨细胞功能障碍,进而影响骨的生长与修复。Ⅵ型胶原蛋白在胚胎期骨形成过程中亦发挥重要作用。在小鼠胚胎模型中,Ⅵ型胶原蛋白表达于骨形态发生早期阶段显著上调,并特异性富集于间充质干细胞向成骨细胞谱系定向分化的关键时期。这些研究不仅深化了对Ⅵ型胶原蛋白在骨骼发育中作用的认识,也为再生医学研究奠定了理论基础[4]。综上所述,深入阐明Ⅵ型胶原蛋白调控成骨细胞功能的多重机制及其在骨骼发育中的时空表达模式,不仅显著拓展了对骨生物学过程的认识,也为发展基于细胞外基质的骨再生修复策略提供了重要的理论依据。见表2。 2.3 Ⅵ型胶原蛋白与骨骼疾病 2.3.1 骨质疏松症中Ⅵ型胶原蛋白的研究进展 骨质疏松症是一种以骨密度降低和骨组织微结构破坏为特征的全身性骨骼疾病。作为细胞外基质的重要组成部分,Ⅵ型胶原蛋白在维持骨组织结构和功能方面起关键作用,Ⅵ型胶原蛋白表达或功能异常与骨质疏松症的发病机制密切相关。在骨质疏松症患者中,Ⅵ型胶原蛋白的表达下调,这种表达下调可能削弱骨基质的力学稳定性,并参与骨密度的下降过程。临床研究观察到,骨质疏松症患者血清中Ⅵ型胶原蛋白降解产物水平显著升高,提示体内Ⅵ型胶原蛋白的降解加速,这与骨质疏松症的进展存在关联[30]。此外,Ⅵ型胶原蛋白缺失可导致成骨细胞功能减弱,进而影响骨基质的合成与矿"
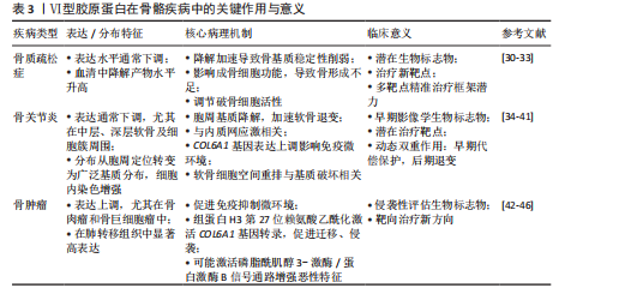
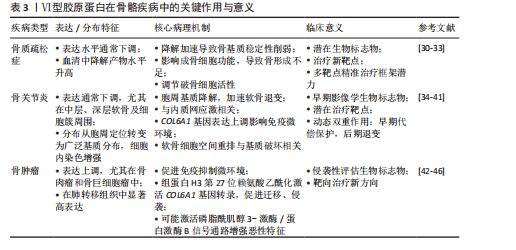
化,最终引起骨密度降低。Ⅵ型胶原蛋白还可通过调控破骨细胞活性,进一步影响骨质疏松的病理进程。具体而言,Ⅵ型胶原蛋白通过调控核因子κB受体活化因子配体与骨保护素的表达比例,影响破骨细胞的分化与活性,从而维持骨稳态[31]。Ⅵ型胶原蛋白缺失还可能削弱成骨细胞的功能,导致骨形成不足,进而加重骨质疏松的病理过程。因此,Ⅵ型胶原蛋白不仅有望作为骨质疏松症的潜在生物标志物,也可能成为未来治疗干预的新靶点。 Ⅵ型胶原蛋白在骨质疏松症治疗中展现出多靶点调控潜力,其编码基因通过不同机制协同调控骨代谢平衡:COL6A2受血清外泌体miR-128-2-5p的表观遗传调控,其表达下调可导致成骨细胞黏附与代谢功能障碍,为绝经后骨质疏松症的干预提供了新靶点[32];而COL6A3可通过增强骨髓间充质干细胞的线粒体自噬能力,显著缓解炎症微环境中的氧化应激损伤,并促进成骨分化[33]。上述机制共同构成一个协同调控网络。因此,靶向COL6A2的miRNA拮抗策略与COL6A3基因过表达技术共同构成潜在的精准治疗框架,但临床转化仍面临组织靶向递送效率低等瓶颈,且需进一步阐明各亚型间的相互作用机制。未来研究应结合单细胞多组学技术,绘制Ⅵ型胶原蛋白在骨组织中的时空表达图谱,并依托3D生物打印骨类器官模型验证多靶点协同干预的有效性,以推动Ⅵ型胶原蛋白发展为骨质疏松治疗的革新性分子靶标群。 2.3.2 骨关节炎中Ⅵ型胶原蛋白的研究进展 近年来,Ⅵ型胶原蛋白在骨关节炎中的作用日益受到关注。在正常关节软骨中,Ⅵ型胶原蛋白主要定位于软骨细胞周围的胞周基质,为软骨细胞提供结构支撑并参与信号调控[34-35]。然而,在骨关节炎软骨中Ⅵ型胶原蛋白表达通常下调,尤以中层、深层区域及软骨细胞簇周围最为显著[35-37];同时,Ⅵ型胶原蛋白分布模式亦发生改变,由胞周定位为主转为更广泛的基质分布,且细胞内免疫染色增强,提示软骨细胞可能代偿性上调Ⅵ型胶原蛋白的合成[35-36]。功能研究表明,Ⅵ型胶原蛋白对软骨具有保护作用:可减轻炎症因子(如白细胞介素1β)诱导的损伤,有效抑制基质降解酶(基质金属蛋白酶3、基质金属蛋白酶9、基质金属蛋白酶13)及一氧化氮的产生,从而延缓软骨基质降解;此外,Ⅵ型胶原蛋白还可促进软骨细胞增殖,并维持基质成分(如Ⅰ型胶原蛋白、聚集蛋白聚糖)的表达[34]。值得注意的是,Ⅵ型胶原蛋白及其构成的胞周基质在骨关节炎进展中发挥复杂作用:随着疾病进展,软骨细胞发生空间重排(由单行排列逐渐演变为双排、小簇乃至大簇),该现象被视为骨关节炎的组织学标志之一;与此同时,以Ⅵ型胶原蛋白为主的胞周基质逐步降解,被认为是骨关节炎的早期病理特征之一[38-39];DANALACHE等[38]通过酶联免疫吸附定量分析发现,Ⅵ型胶原蛋白的信号强度与蛋白含量均显著降低,且该变化与软骨细胞的空间组织模式密切相关[40];研究表明,基质金属蛋白酶(基质金属蛋白酶2、基质金属蛋白酶3、基质金属蛋白酶7)在骨关节炎早期即可降解Ⅵ型胶原蛋白,导致胞周基质结构破坏,加速软骨退变[38-39],Ⅵ型胶原蛋白缺失被认为是关节软骨退化的关键因素,可能通过削弱细胞-基质相互作用而促进骨关节炎进展[40];此外,Ⅵ型胶原蛋白的异常还与内质网应激密切相关;骨关节炎软骨细胞中内质网应激标志物(如葡萄糖调节蛋白78、Bcl-2相关抗凋亡基因1)表达上调,提示细胞处于应激状态[36]。XU等[41]通过多组学数据分析与实验验证发现,在骨关节炎患者亚软骨骨组织及破骨细胞分化过程中COL6A1表达显著上调,且COL6A1表达水平与局部免疫微环境变化密切相关。综上所述,Ⅵ型胶原蛋白是骨关节炎病理过程的关键参与者和潜在生物标志物,COL6A1表达与分布的改变、功能丧失以及胞周基质结构的破坏,均深度参与疾病的发生与发展,因而被视为潜在的治疗靶点[40]。基于现有证据,Ⅵ型胶原蛋白在骨关节炎中呈现动态的双重作用:早期表达上调可能为一种代偿性保护反应,旨在维持软骨稳态,然而随病程进展,Ⅵ型胶原蛋白降解及胞周基质结构破坏,尤其是伴随软骨细胞空间重排,构成软骨退变的核心病理环节。值得注意的是,Ⅵ型胶原蛋白含量变化与软骨细胞组织模式的关联性,使它有望成为骨关节炎早期的潜在影像学生物标志物。更重要的是,Ⅵ型胶原蛋白在调控细胞微环境中的关键作用,使它成为干预软骨退变、延缓骨关节炎进展极具前景的治疗靶点,亟需深入解析Ⅵ型胶原蛋白调控机制并开发靶向干预策略。 2.3.3 骨肿瘤中Ⅵ型胶原蛋白的研究进展 近年研究表明,Ⅵ型胶原蛋白在骨肿瘤的发生与发展中发挥重要作用,Ⅵ型胶原蛋白表达特征与肿瘤微环境及恶性生物学行为密切相关。Ⅵ型胶原蛋白及其蛋白片段在肿瘤相关成纤维细胞中高表达,可通过与免疫细胞表面受体(如白细胞相关免疫球蛋白样受体1)结合,显著抑制T细胞、NK细胞、巨噬细胞及树突状细胞的功能,从而促进免疫抑制性微环境的形成,该机制已被多项研究证实,阻断此相互作用可增强抗肿瘤免疫应答[42]。泛癌种生物信息学分析亦表明,COL6A1、COL6A2与COL6A3的表达水平与肿瘤免疫亚型及免疫细胞浸润程度密切相关[43]。在骨肉瘤中,Ⅵ型胶原蛋白α1链普遍上调,尤其在肺转移灶中显著高表达,且与患者不良预后相关。机制研究表明,组蛋白H3第27位赖氨酸(H3K27)乙酰化可激活COL6A1的转录,促进骨肉瘤细胞的迁移与侵袭,该过程依赖于Ⅵ型胶原蛋白α1链与细胞因子信号转导抑制因子5的相互作用,进而抑制信号转导及转录激活因子1的表达与活化。此外,骨肉瘤细胞分泌的外泌体富含Ⅵ型胶原蛋白α1链,可通过激活转化生长因子β/COL6A1信号轴,诱导正常成纤维细胞向肿瘤相关成纤维细胞转化,从而构建促肿瘤微环境,加剧骨肉瘤细胞的侵袭与转移[44]。值得注意的是,Ⅵ型胶原蛋白的异常表达不仅见于骨肉瘤,在其他类型骨肿瘤中亦具有显著特征。例如,在骨巨细胞瘤等侵袭性骨肿瘤中,Ⅵ型胶原蛋白表达水平显著升高,且与肿瘤的侵袭性及转移潜能呈正相关[45]。进一步研究表明,Ⅵ型胶原蛋白上调可能通过激活磷脂酰肌醇3-激酶/蛋白激酶B信号通路,增强肿瘤细胞的迁移、侵袭等恶性表型,从而促进骨肿瘤进展[46]。综上,Ⅵ型胶原蛋白可作为评估骨肿瘤侵袭性的潜在生物标志物,其调控网络及相关信号通路亦为骨肿瘤的靶向治疗提供了新策略。见表3。"

2.4 未来研究方向 2.4.1 Ⅵ型胶原蛋白在组织工程研究中的应用 随着生物技术的快速发展,新兴技术为Ⅵ型胶原蛋白研究提供了重要机遇:①基因工程技术可用于制备重组Ⅵ型胶原蛋白,不仅可提高产量,还能维持结构完整性与生物活性,并在一定程度上模拟天然合成及翻译后修饰过程[47];在组织修复与细胞保护方面,Ⅵ型胶原蛋白作为细胞外基质的关键组分,凭借其独特结构与多重生物学功能展现出广阔应用前景。例如,Ⅵ型胶原蛋白以串珠状微纤维形式广泛分布于皮肤、肺、血管及关节软骨等多种组织中,可将细胞锚定于基质,并介导生化与力学信号的传递[48-49];此外,Ⅵ型胶原蛋白可用于模拟天然骨的有机基质成分,从而增强人骨髓间充质干细胞的成骨分化能力及支架的力学性能。研究表明,在工程化骨移植物中引入Ⅵ型胶原蛋白可促进成骨相关基因表达,进而提升骨愈合效率[24]。此外,Ⅵ型胶原蛋白还具有显著的细胞保护作用,包括抑制凋亡、减轻氧化损伤、调控自噬、促进细胞分化及维持干细胞干性等[49]。研究报道,Ⅵ型胶原蛋白缺失可导致巨核细胞自噬受损、内质网应激增强及凋亡增加;而通过激活自噬(如采用饥饿或雷帕霉素处理)可恢复自噬流,缓解内质网应激与细胞凋亡,从而改善细胞功能[50];在仿生材料领域,Ⅵ型胶原蛋白的微观结构与多级组装特征为仿生设计提供了理论基础;将Ⅵ型胶原蛋白整合至支架中可显著改善材料的力学性能与生物活性,对骨缺损修复具有重要意义[24]。更重要的是,Ⅵ型胶原蛋白在维持组织力学性能、促进细胞存活及功能恢复方面具有独特优势,适用于软骨、皮肤和神经等多种组织的再生修复[48-49];通过将Ⅵ型胶原蛋白与生物活性分子或其他功能材料复合,可进一步拓展其生物学功能与应用范围[51-53]。 未来,Ⅵ型胶原蛋白的多层次结构解析为新型仿生支架材料的开发奠定了理论基础,但在大规模制备、功能精准调控及多材料协同应用等方面仍需深入研究[48-49,52],上述技术的发展不仅为基础研究提供了新工具,也为临床转化创造了条件;未来研究应聚焦于Ⅵ型胶原蛋白相关疾病的靶向干预策略。Ⅵ型胶原蛋白凭借独特的微观结构、优异的生物力学特性及多重生物学功能,在组织工程支架构建中凸显核心价值[54],为临床转化指明方向。然而,要实现从基础研究到临床规模化应用的转化,亟需在重组蛋白的高效制备工艺、功能化修饰策略及多材料协同作用机制等关键环节取得突破。未来研究应着力解析Ⅵ型胶原蛋白的结构-功能关系图谱,并构建智能化递送系统,以加速Ⅵ型胶原蛋白基生物材料在再生医学领域的临床转化进程。 2.4.2 Ⅵ型胶原蛋白作为骨骼生物标志物的潜力 鉴于Ⅵ型胶原蛋白在骨骼系统细胞外基质中的关键作用及其与多种骨骼疾病的密切关联,它已成为骨骼系统疾病生物标志物研究的一个热点。 研究表明,与Ⅵ型胶原蛋白降解和合成相关的特定片段,例如基质金属蛋白酶降解片段(C6M)、Ⅵ型原胶原C端结构域(PRO-C6)以及N端球状结构域表位,均可在血清中被检测到,并能动态反映骨骼肌肉系统的病理生理变化。值得注意的是,这些片段的功能具有特异性:N端球状结构域表位水平在年轻男性中与肌肉质量显著相关,而基质金属蛋白酶降解片段则与肌肉再生过程特异性关联,这提示它们可作为评估肌肉动态平衡的潜在标志物[55]。在遗传性骨病领域,Ⅵ型胶原蛋白相关标志物在疾病分型与治疗监测方面展现出重要价值。例如,在成骨不全患者中,接受特立帕肽治疗后,血清Ⅵ型胶原蛋白标志物水平显著变化揭示了骨基质重塑过程中的生物力学响应差异,且不同亚型患者呈现出独特的标志物变化模式,这为实施精准治疗提供了新依据[30]。这些标志物的应用已从内脏器官纤维化研究拓展至骨骼系统。例如,Ⅵ型原胶原C端结构域和基质金属蛋白酶降解片段在系统性硬化症等疾病中表现出优异的诊断效能(受试者工作特征曲线下面积大于0.90),其水平变化不仅能有效区分健康与疾病状态,还可用于评估骨骼肌纤维化程度及疾病进展[56-57]。 此外,Ⅵ型胶原蛋白的表达异常还与关节挛缩、骨骼发育缺陷等先天性表型直接相关,这为Ⅵ型胶原蛋白在先天性骨骼疾病早期诊断中的应用提供了分子基础[1,11,58]。 综上所述,关于Ⅵ型胶原蛋白相关生物标志物的研究已从基础机制探索逐步向临床转化阶段迈进。现有证据表明,血清Ⅵ型胶原蛋白标志物不仅能反映骨骼肌肉系统的结构完整性,还可动态监测治疗干预所引发的基质重塑过程,因而在疾病诊断、分型及疗效评估方面均展现出多重应用价值。然而,该领域研究目前仍存在样本量有限、检测方法标准化不足等局限性。未来研究需通过大规模多中心队列验证这些标志物的临床敏感性与特异性,建立Ⅵ型胶原蛋白相关生物标志物与影像学、病理学改变相关联的综合评价体系,并积极探索在骨骼组织工程与再生治疗中的动态监测价值。实现这些突破,将有力推动Ⅵ型胶原蛋白相关标志物在骨骼系统疾病精准医疗实践中的重要工具。 2.4.3 研发针对Ⅵ型胶原蛋白的治疗策略 针对Ⅵ型胶原蛋白相关疾病的治疗策略研究正从传统的对症支持治疗迅速转向分子靶向与再生医学方向,目前主要聚焦于基因靶向、RNA干预、细胞移植及下游信号通路调控等前沿领域。在基因与RNA靶向治疗方面,等位基因特异性沉默策略利用小干扰RNA或gapmer型反义寡核苷酸,可选择性沉默致病突变等位基因,同时保留野生型等位基因的表达,从而有效恢复Ⅵ型胶原蛋白的正常分泌与功能[59-61]。此外,针对Ⅵ型胶原蛋白α1链常见的深内含子突变,反义寡核苷酸介导的外显子跳跃技术可特异性诱导异常外显子跳过,有效恢复mRNA的正常剪接,进而重建具有功能的Ⅵ型胶原蛋白微纤维[61-62]。在细胞替代与再生治疗领域,系统性移植诱导多能干细胞来源的间充质干细胞已在小鼠模型中证实可补充Ⅵ型胶原蛋白,显著改善肌肉结构与功能,并增强肌肉再生能力[63]。值得注意的是,针对Ⅵ型胶原蛋白缺失所引发的下游病理改变,调控关键信号通路及代谢过程亦展现出良好前景。研究利用Ⅵ型胶原蛋白α2链基因敲除小鼠模型证实,Ⅵ型胶原蛋白缺失可导致转化生长因子β生物利用度失调,因此调控该信号通路有望缓解相关肌肉病理改变[64];同时,Ⅵ型胶原蛋白缺陷所致的线粒体功能障碍与细胞凋亡,可通过线粒体通透性转换孔抑制剂(如环孢素A)部分逆转[65]。另外,鉴于Ⅵ型胶原蛋白参与调控自噬以维持细胞稳态,增强自噬活性或保护细胞免受凋亡及氧化应激损伤的药物亦被视为潜在治疗策略[66]。此外,整合基因共表达网络分析与代谢建模研究已识别出关键枢纽基因及代谢物,为药物再利用与新靶点开发提供了重要依据[11,67]。 值得注意的是,部分已上市药物可通过调控Ⅵ型胶原蛋白相关信号通路,间接影响Ⅵ型胶原蛋白表达或功能,展现出药物再利用的潜力。例如,雷帕霉素通过激活自噬通路,可部分逆转Ⅵ型胶原蛋白缺陷所致的内质网应激与细胞死亡[50, 66];天然多酚紫檀芪(pterostilbene)亦被证实可通过增强自噬,改善Ⅵ型胶原蛋白缺失小鼠模型的病理表型[68]。此外,转化生长因子β信号通路抑制剂在纤维化模型中可下调COL6A1、COL6A2和COL6A3基因表达,抑制成纤维细胞活化及细胞外基质异常沉积[69],提示其在Ⅵ型胶原蛋白高表达相关的骨肿瘤或纤维性骨病中具有潜在应用价值。综上所述,等位基因特异性沉默、反义寡核苷酸介导的剪接修饰、干细胞移植及下游信号通路调控等策略在细胞及动物模型中均显示出显著疗效,为未来临床转化奠定了坚实基础。"

| [1] DI MARTINO A, CESCON M, D’AGOSTINO C, et al. Collagen VI in the Musculoskeletal System. Int J Mol Sci. 2023;24(6):5095. [2] KOMORI T, JI Y, PHAM H, et al. Type VI Collagen Regulates Endochondral Ossification in the Temporomandibular Joint. JBMR Plus. 2022;6(5): e10617. [3] NISSEN NI, KARSDAL M, WILLUMSEN N. Collagens and Cancer associated fibroblasts in the reactive stroma and its relation to Cancer biology. J Exp Clin Cancer Res. 2019;38(1):115. [4] AHMED S, NOWLAN NC. Initiation and emerging complexity of the collagen network during prenatal skeletal development. Eur Cell Mater. 2020;39:136-155. [5] WANG PP, DING SY, SUN YY, et al. MYCT1 Inhibits the Adhesion and Migration of Laryngeal Cancer Cells Potentially Through Repressing Collagen VI. Front Oncol. 2021;10:564733. [6] GREGORY CA, MA J, LOMELI S. The coordinated activities of collagen VI and XII in maintenance of tissue structure, function and repair: evidence for a physical interaction. Front Mol Biosci. 2024; 11:1376091. [7] SOLOMON-DEGEFA H, GEBAUER JM, JEFFRIES CM, et al. Structure of a collagen VI α3 chain VWA domain array: adaptability and functional implications of myopathy causing mutations. J Biol Chem. 2020;295(36):12755-12771. [8] ZANOTTI S, MAGRI F, SALANI S, et al. Extracellular Matrix Disorganization and Sarcolemmal Alterations in COL6-Related Myopathy Patients with New Variants of COL6 Genes. Int J Mol Sci. 2023;24(6):5551. [9] NALLANTHIGHAL S, HEISERMAN JP, CHEON DJ. Collagen Type XI Alpha 1 (COL11A1): A Novel Biomarker and a Key Player in Cancer. Cancers (Basel). 2021;13(5):935. [10] MERENESS JA, MARIANI TJ. The critical role of collagen VI in lung development and chronic lung disease. Matrix Biol Plus. 2021;10:100058. [11] LAMANDÉ SR, BATEMAN JF. Collagen VI disorders: Insights on form and function in the extracellular matrix and beyond. Matrix Biol. 2018;71-72:348-367. [12] ANTONIEL M, TRAINA F, MERLINI L, et al. Tendon Extracellular Matrix Remodeling and Defective Cell Polarization in the Presence of Collagen VI Mutations. Cells. 2020;9(2):409. [13] LEIPHART RJ, PHAM H, HARVEY T, et al. Coordinate roles for collagen VI and biglycan in regulating tendon collagen fibril structure and function. Matrix Biol Plus. 2021;13:100099. [14] CENNI V, SABATELLI P, DI MARTINO A, et al. Collagen VI Deficiency Impairs Tendon Fibroblasts Mechanoresponse in Ullrich Congenital Muscular Dystrophy. Cells. 2024;13(5):378. [15] WANG X, JIA Q, YU L, et al. Filamin B knockdown impairs differentiation and function in mouse pre-osteoblasts via aberrant transcription and alternative splicing. Heliyon. 2024;10(20):e39334. [16] IZU Y, EZURA Y, KOCH M, et al. Collagens VI and XII form complexes mediating osteoblast interactions during osteogenesis. Cell Tissue Res. 2016;364(3):623-635. [17] KOMORI T, PHAM H, JANI P, et al. The Role of Type VI Collagen in Alveolar Bone. Int J Mol Sci. 2022;23(22):14347. [18] IZU Y, EZURA Y, MIZOGUCHI F, et al. Type VI collagen deficiency induces osteopenia with distortion of osteoblastic cell morphology. Tissue Cell. 2012;44(1):1-6. [19] CHRISTENSEN SE, COLES JM, ZELENSKI NA, et al. Altered trabecular bone structure and delayed cartilage degeneration in the knees of collagen VI null mice. PLoS One. 2012;7(3):e33397. [20] PHAM HT, KRAM V, DAR QA, et al. Collagen VIα2 chain deficiency causes trabecular bone loss by potentially promoting osteoclast differentiation through enhanced TNFα signaling. Sci Rep. 2020; 10(1):13749. [21] KOMORI T, KRAM V, PERRY S, et al. Type VI Collagen Deficiency Causes Enhanced Periodontal Tissue Destruction. J Dent Res. 2024;103(9):878-888. [22] MARGIOTTA A. Coupling of Intracellular Calcium Homeostasis and Formation and Secretion of Matrix Vesicles: Their Role in the Mechanism of Biomineralization. Cells. 2025;14(10):733. [23] HASEGAWA T, YAMAMOTO T, TSUCHIYA E, et al. Ultrastructural and biochemical aspects of matrix vesicle-mediated mineralization. Jpn Dent Sci Rev. 2017;53(2):34-45. [24] MONDRAGÓN E, COWDIN M, TARABALLI F, et al. Mimicking the Organic and Inorganic Composition of Anabolic Bone Enhances Human Mesenchymal Stem Cell Osteoinduction and Scaffold Mechanical Properties. Front Bioeng Biotechnol. 2020;8:753. [25] NUDELMAN F, PIETERSE K, GEORGE A, et al. The role of collagen in bone apatite formation in the presence of hydroxyapatite nucleation inhibitors. Nat Mater. 2010;9(12):1004-1009. [26] DU T, NIU X, HOU S, et al. Apatite minerals derived from collagen phosphorylation modification induce the hierarchical intrafibrillar mineralization of collagen fibers. J Biomed Mater Res A. 2019; 107(11):2403-2413. [27] OGURA H, NAKAMURA T, ISHII T, et al. Mechanical stress-induced FGF-2 promotes proliferation and consequently induces osteoblast differentiation in mesenchymal stem cells. Biochem Biophys Res Commun. 2023;684:149145. [28] ZHANG N, JIANG H, BAI Y, et al. The molecular mechanism study of insulin on proliferation and differentiation of osteoblasts under high glucose conditions. Cell Biochem Funct. 2019;37(5):385-394. [29] KOHARA Y, SOETA S, IZU Y, et al. Distribution of type VI collagen in association with osteoblast lineages in the groove of Ranvier during rat postnatal development. Ann Anat. 2016;208:58-68. [30] NICOL L, SRIKANTH P, HENRIKSEN K, et al. Widespread disturbance in extracellular matrix collagen biomarker responses to teriparatide therapy in osteogenesis imperfecta. Bone. 2021; 142:115703. [31] LIU K, LIU Y, XU Y, et al. Asperosaponin VI protects against bone destructions in collagen induced arthritis by inhibiting osteoclastogenesis. Phytomedicine. 2019;63:153006. [32] LU L, WANG L, WANG H, et al. Regulatory role of miR-128-2-5p in serum exosomes on COL6A2 expression and postmenopausal osteoporosis. Hum Mol Genet. 2025;34(7):563-576. [33] WANG K, PENG X, ZHANG R, et al. COL6A3 enhances the osteogenic differentiation potential of BMSCs by promoting mitophagy in the osteoporotic microenvironment. Mol Biol Rep. 2024;51(1):206. [34] CHU WC, ZHANG S, SNG TJ, et al. Distribution of pericellular matrix molecules in the temporomandibular joint and their chondroprotective effects against inflammation. Int J Oral Sci. 2017;9(1):43-52. [35] PULLIG O, WESELOH G, SWOBODA B. Expression of type VI collagen in normal and osteoarthritic human cartilage. Osteoarthritis Cartilage. 1999; 7(2):191-202. [36] NUGENT AE, SPEICHER DM, GRADISAR I, et al. Advanced osteoarthritis in humans is associated with altered collagen VI expression and upregulation of ER-stress markers Grp78 and bag-1. J Histochem Cytochem. 2009;57(10):923-931. [37] MCDEVITT CA, PAHL JA, AYAD S, et al. Experimental osteoarthritic articular cartilage is enriched in guanidine soluble type VI collagen. Biochem Biophys Res Commun. 1988;157(1):250-255. [38] DANALACHE M, ERLER AL, WOLFGART JM, et al. Biochemical changes of the pericellular matrix and spatial chondrocyte organization-Two highly interconnected hallmarks of osteoarthritis. J Orthop Res. 2020;38(10):2170-2180. [39] DANALACHE M, UMRATH F, RIESTER R, et al. Proteolysis of the pericellular matrix: Pinpointing the role and involvement of matrix metalloproteinases in early osteoarthritic remodeling. Acta Biomater. 2024;181:297-307. [40] YOTSUYA M, BERTAGNA AE, HASAN N, et al. Neuron/Glial Antigen 2-Type VI Collagen Interactions During Murine Temporomandibular Joint Osteoarthritis. Sci Rep. 2019;9(1):56. [41] XU Q, FENG G, TANG Z, et al. Mechanistic study of COL6A1-mediated subchondral bone remodeling in osteoarthritis via the EPAC/RAP1 axis. FASEB J. 2025;39(6):e70473. [42] FLIES DB, LANGERMANN S, JENSEN C, et al. Regulation of tumor immunity and immunotherapy by the tumor collagen extracellular matrix. Front Immunol. 2023;14: 1199513. [43] LI X, LI Z, GU S, et al. A pan-cancer analysis of collagen VI family on prognosis, tumor microenvironment, and its potential therapeutic effect. BMC Bioinformatics. 2022;23(1):390. [44] ZHANG Y, LIU Z, YANG X, et al. H3K27 acetylation activated-COL6A1 promotes osteosarcoma lung metastasis by repressing STAT1 and activating pulmonary cancer-associated fibroblasts. Theranostics. 2021;11(3):1473-1492. [45] SUGITA S, TAKENAMI T, KIDO T, et al. Diagnostic utility of CSF1 immunohistochemistry in tenosynovial giant cell tumor for differentiating from giant cell-rich tumors and tumor-like lesions of bone and soft tissue. Diagn Pathol. 2022;17(1):88. [46] GUO HL, CHEN G, SONG ZL, et al. COL6A3 promotes cellular malignancy of osteosarcoma by activating the PI3K/AKT pathway. Rev Assoc Med Bras (1992). 2020;66(6):740-745. [47] FERTALA A. Three Decades of Research on Recombinant Collagens: Reinventing the Wheel or Developing New Biomedical Products? Bioengineering (Basel). 2020;7(4):155. [48] GODWIN ARF, STARBORG T, SHERRATT MJ, et al. Defining the hierarchical organisation of collagen VI microfibrils at nanometre to micrometre length scales. Acta Biomater. 2017;52:21-32. [49] CESCON M, GATTAZZO F, CHEN P, et al. Collagen VI at a glance. J Cell Sci. 2015;128(19):3525-3531. [50] ABBONANTE V, MALARA A, CHRISAM M, et al. Lack of COL6/collagen VI causes megakaryocyte dysfunction by impairing autophagy and inducing apoptosis. Autophagy. 2023;19(3):984-999. [51] LIU C. Application of marine collagen for stem-cell-based therapy and tissue regeneration (Review). Med Int (Lond). 2021;1(3):6. [52] ULLAH S, ZAINOL I. Fabrication and applications of biofunctional collagen biomaterials in tissue engineering. Int J Biol Macromol. 2025;298: 139952. [53] DONG C, LV Y. Application of Collagen Scaffold in Tissue Engineering: Recent Advances and New Perspectives. Polymers (Basel). 2016;8(2):42. [54] 侯建明,李琦.生物活性支架材料修复骨质疏松性骨缺损[J].中国组织工程研究,2023,27(21): 3423-3429. [55] NEDERGAARD A, SUN S, KARSDAL MA, et al. Type VI collagen turnover-related peptides-novel serological biomarkers of muscle mass and anabolic response to loading in young men. J Cachexia Sarcopenia Muscle. 2013;4(4):267-275. [56] JUHL P, BAY-JENSEN AC, KARSDAL M, et al. Serum biomarkers of collagen turnover as potential diagnostic tools in diffuse systemic sclerosis: A cross-sectional study. PLoS One. 2018;13(12):e0207324. [57] WILLIAMS L, LAYTON T, YANG N, et al. Collagen VI as a driver and disease biomarker in human fibrosis. FEBS J. 2022;289(13):3603-3629. [58] KWONG AK, ZHANG Y, HO RS, et al. Collagen VI-related myopathies: clinical variability, phenotype-genotype correlation and exploratory transcriptome study. Neuromuscul Disord. 2023; 33(5):371-381. [59] BRULL A, SARATHY A, BOLDUC V, et al. Optimized allele-specific silencing of the dominant-negative COL6A1 G293R substitution causing collagen VI-related dystrophy. Mol Ther Nucleic Acids. 2024;35(2):102178. [60] MARROSU E, ALA P, MUNTONI F, et al. Gapmer Antisense Oligonucleotides Suppress the Mutant Allele of COL6A3 and Restore Functional Protein in Ullrich Muscular Dystrophy. Mol Ther Nucleic Acids. 2017;8:416-427. [61] GUALANDI F, MANZATI E, SABATELLI P, et al. Antisense-induced messenger depletion corrects a COL6A2 dominant mutation in Ullrich myopathy. Hum Gene Ther. 2012;23(12):1313-1318. [62] AGUTI S, BOLDUC V, ALA P, et al. Exon-Skipping Oligonucleotides Restore Functional Collagen VI by Correcting a Common COL6A1 Mutation in Ullrich CMD. Mol Ther Nucleic Acids. 2020; 21:205-216. [63] HARADA A, GOTO M, KATO A, et al. Systemic Supplementation of Collagen VI by Neonatal Transplantation of iPSC-Derived MSCs Improves Histological Phenotype and Function of Col6-Deficient Model Mice. Front Cell Dev Biol. 2021; 9:790341. [64] MOHASSEL P, HEARN H, ROONEY J, et al. Collagen type VI regulates TGF-β bioavailability in skeletal muscle in mice. J Clin Invest. 2025; 135(9):e173354. [65] IRWIN WA, BERGAMIN N, SABATELLI P, et al. Mitochondrial dysfunction and apoptosis in myopathic mice with collagen VI deficiency. Nat Genet. 2003;35(4):367-371. [66] CASTAGNARO S, GAMBAROTTO L, CESCON M, et al. Autophagy in the mesh of collagen VI. Matrix Biol. 2021;100-101:162-172. [67] CEYHAN AB, KAYNAR A, ALTAY O, et al. Identifying Hub Genes and Metabolic Pathways in Collagen VI-Related Dystrophies: A Roadmap to Therapeutic Intervention. Biomolecules. 2024;14(11):1376. [68] METTI S, GAMBAROTTO L, CHRISAM M, et al. The Polyphenol Pterostilbene Ameliorates the Myopathic Phenotype of Collagen VI Deficient Mice via Autophagy Induction. Front Cell Dev Biol. 2020;8:580933. [69] WILLIAMS LM, MCCANN FE, CABRITA MA, et al. Identifying collagen VI as a target of fibrotic diseases regulated by CREBBP/EP300. Proc Natl Acad Sci U S A. 2020;117(34):20753-20763. |
| [1] | Chen Huiting, Zeng Weiquan, Zhou Jianhong, Wang Jie, Zhuang Congying, Chen Peiyou, Liang Zeqian, Deng Weiming. Tail anchoring technique of vertebroplasty in treatment of osteoporotic vertebral compression fractures with intravertebral cleft: a finite element analysis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2145-2152. |
| [2] | Zeng Xuan, Weng Rui, Ye Shicheng, Tang Jiadong, Mo Ling, Li Wenchao. Two lumbar rotary manipulation techniques in treating lumbar disc herniation: a finite element analysis of biomechanical differences [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2153-2161. |
| [3] | Cheng Qisheng, Julaiti·Maitirouzi, Xiao Yang, Zhang Chenwei, Paerhati·Rexiti. Finite element analysis of novel variable-diameter screws in modified cortical bone trajectory of lumbar vertebrae [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2162-2171. |
| [4] | Liu Wenlong, Dong Lei, Xiao Zhengzheng, Nie Yu. Finite element analysis of tibial prosthesis loosening after fixed-bearing unicompartmental knee arthroplasty for osteoporosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2191-2198. |
| [5] | Chen Long, Wang Xiaozhen, Xi Jintao, Lu Qilin. Biomechanical performance of short-segment screw fixation combined with expandable polyetheretherketone vertebral body replacement in osteoporotic vertebrae [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2226-2235. |
| [6] | Hu Xiongke, Liu Shaohua, Tan Qian, Liu Kun, Zhu Guanghui. Shikonin intervention with bone marrow mesenchymal stem cells improves microstructure of femur in aged mice [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1609-1615. |
| [7] | Wen Guangwei, Zhen Yinghao, Zheng Taikeng, Zhou Shuyi, Mo Guoye, Zhou Tengpeng, Li Haishan, Lai Yiyi. Effects and mechanisms of isoginkgetin on osteoclastogenesis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(6): 1348-1358. |
| [8] | Wu Zhilin, , He Qin, Wang Pingxi, Shi Xian, Yuan Song, Zhang Jun, Wang Hao . DYRK2: a novel therapeutic target for rheumatoid arthritis combined with osteoporosis based on East Asian and European populations [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(6): 1569-1579. |
| [9] | Zhang Haiwen, Zhang Xian, Xu Taichuan, Li Chao. Bibliometric and visual analysis of the research status and trends of senescence in osteoporosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(6): 1580-1591. |
| [10] | Huang Jie, Zeng Hao, Wang Wenchi, Lyu Zhucheng, Cui Wei. Visualization analysis of literature on the effect of lipid metabolism on osteoporosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(6): 1558-1568. |
| [11] | Yang Zhijie, Zhao Rui, Yang Haolin, Li Xiaoyun, Li Yangbo, Huang Jiachun, Lin Yanping, Wan Lei, HuangHongxing. Postmenopausal osteoporosis: predictive values of muscle mass, grip strength, and appendicular skeletal muscle index [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(5): 1073-1080. |
| [12] | Zhou Jian, Zhang Tao, Zhou Weili, Zhao Xingcheng, Wang Jun, Shen Jie, Qian Li, Lu Ming. Effects of resistance training on quadriceps mass and knee joint function in patients with osteoporosis and sarcopenia [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(5): 1081-1088. |
| [13] | Cao Wenqi, Feng Xiuzhi, Zhao Yi, Wang Zhimin, Chen Yiran, Yang Xiao, Ren Yanling. Effect of macrophage polarization on osteogenesis-angiogenesis coupling in type 2 diabetic osteoporosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(4): 917-925. |
| [14] | Zeng Hao, Sun Pengcheng, Chai Yuan, Huang Yourong, Zhang Chi, Zhang Xiaoyun. Association between thyroid function and osteoporosis: genome-wide data analysis of European populations [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(4): 1019-1027. |
| [15] | Zhu Lifeng, Wang Wenchi, Liu Qiang, Cui Xianqin, Zhang Zhenhao, Huang Jie, Lyu Zhucheng, Wang Leihang, Cui Wei. Molecular mechanism of icariin in prevention and treatment of osteoporosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(35): 9248-9257. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||