Chinese Journal of Tissue Engineering Research ›› 2026, Vol. 30 ›› Issue (35): 9248-9257.doi: 10.12307/2026.283
Previous Articles Next Articles
Molecular mechanism of icariin in prevention and treatment of osteoporosis
Zhu Lifeng1, Wang Wenchi1, Liu Qiang2, Cui Xianqin1, Zhang Zhenhao1, Huang Jie1, Lyu Zhucheng1, Wang Leihang1, Cui Wei3
- 1Guangxi University of Chinese Medicine, Nanning 530000, Guangxi Zhuang Autonomous Region, China; 2Jiangxi University of Chinese Medicine, Nanchang 330000, Jiangxi Province, China; 3Affiliated Ruikang Hospital of Guangxi University of Chinese Medicine, Nanning 530000, Guangxi Zhuang Autonomous Region, China
-
Received:2025-09-10Revised:2025-12-03Online:2026-12-18Published:2026-04-29 -
Contact:Cui Wei, MD, Chief physician, Professor, Master’s supervisor, Affiliated Ruikang Hospital of Guangxi University of Chinese Medicine, Nanning 530000, Guangxi Zhuang Autonomous Region, China -
About author:Zhu Lifeng, MS candidate, Guangxi University of Chinese Medicine, Nanning 530000, Guangxi Zhuang Autonomous Region, China -
Supported by:Guizhou School of Traditional Chinese Medicine Inheritance and Innovation Team of Guangxi University of Chinese Medicine, No. 2022A004 (to CW); Guangxi Zhuang Autonomous Region Administration of Traditional Chinese Medicine Self-funded Scientific Research Project, No. 20210579 (to CW); Chinese Ethnic Medicine Association Project, No. 2020MZ-040701 (to CW)
CLC Number:
Cite this article
Zhu Lifeng, Wang Wenchi, Liu Qiang, Cui Xianqin, Zhang Zhenhao, Huang Jie, Lyu Zhucheng, Wang Leihang, Cui Wei. Molecular mechanism of icariin in prevention and treatment of osteoporosis[J]. Chinese Journal of Tissue Engineering Research, 2026, 30(35): 9248-9257.
share this article
Add to citation manager EndNote|Reference Manager|ProCite|BibTeX|RefWorks

2.1 淫羊藿苷抗骨质疏松症的研究时间线 自1997年,SHA等[15]首次成功从淫羊藿中分离提纯出淫羊藿苷,为其后续研究奠定了基础。此后,针对淫羊藿苷在骨质疏松症防治中的研究逐步深化。2005年,XIAO团队[16]发现,淫羊藿苷能在体外显著促进大鼠成骨细胞中骨桥蛋白mRNA和Ⅰ型胶原表达。2007年,CHEN等[17]揭示了淫羊藿苷在抑制核因子κB受体活化因子配体和巨噬细胞集落刺激因子诱导的小鼠骨髓破骨细胞形成方面的作用。2010年,MOK等[18]研究进一步证明淫羊藿苷能防止雌激素缺乏引起的骨质流失,淫羊藿苷可能通过不依赖配体的方式激活内质网而在骨中发挥合成代谢作用。随着研究的深入,淫羊藿苷的信号通路机制也逐渐清晰。2012年,LIANG等[19]证实淫羊藿苷通过hFOB 1.19人成骨细胞系中的骨形态发生蛋白2/Smad4信号转导通路促进骨形成。2016年,ZHANG等[20]发现淫羊藿苷通过雌激素受体介导途径刺激原代成骨细胞的分化并抑制其向脂肪细胞分化。2019年,LIANG等[21]的研究表明,淫羊藿苷可通过激活自噬促进骨髓基质细胞的成骨分化,有效防止并卵巢切除小鼠模型的骨质流失。自2019年以来,围绕淫羊藿苷防治骨质疏松症功效及其作用机制的研究仍在持续深入拓展。淫羊藿苷抗骨质疏松症的研究时间线见图4。 2.2 淫羊藿苷防治骨质疏松症的相关性研究 目前已有大量文献从骨组织形态学、生物力学、骨组织生化指标等方面证实了淫羊藿苷的抗骨质疏松作用[22-24],鉴于淫羊藿苷在骨质疏松防治中展现的独特优势,当前研究已聚焦于淫羊藿苷作用机制的深度解析,使之成为该领域的重要研究方向。淫羊藿苷的作用机制主要涉及对骨髓间充质干细胞、成骨细胞及破骨细胞分化与功能的调控。 2.2.1 淫羊藿苷对骨髓间充质干细胞的影响 骨髓间充质干细胞是起源于中胚层的一类非造血源性干细胞群体,具有长期自我更新能力及分化潜能[25],在特定信号调控下,该类细胞可实现向骨细胞、软骨细胞、脂肪细胞、肌细胞、神经细胞及心肌细胞等的定向转化[26]。现代药理学研究证实,淫羊藿苷可通过调控相关信号通路诱导骨髓间充质干细胞向成骨细胞分化,从而在骨代谢平衡调控中发挥干预作用,为骨质疏松症的防治提供了新的作用靶点。 HAN等[27]研究表明,经淫羊藿苷干预后,大鼠骨髓间充质干细胞中TAZ表达激活,成骨指标碱性磷酸酶、Runt相关转录因子2表达"

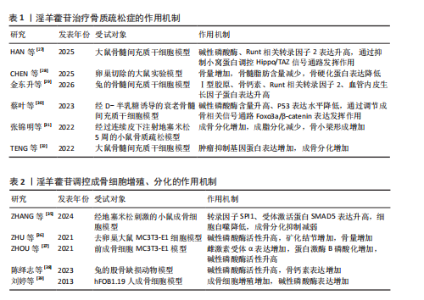
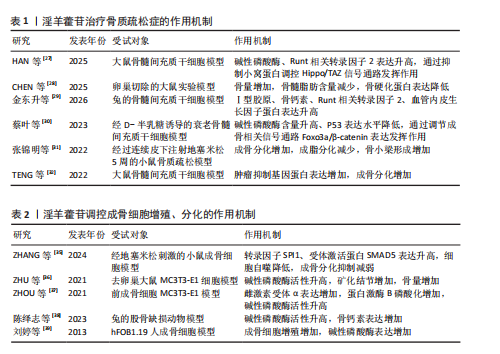
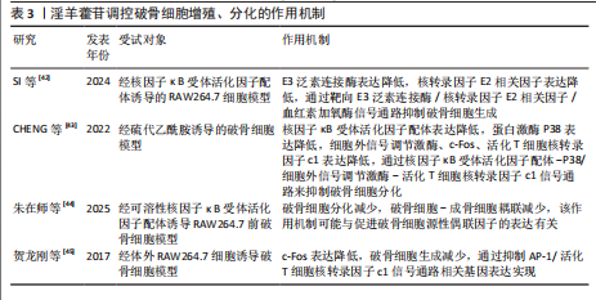
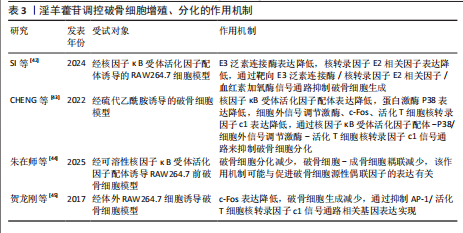
升高,并且最佳干预浓度为1×10-7 mol/L;沉默小窝蛋白显著促进骨髓间充质干细胞的成骨分化,淫羊藿苷通过抑制小窝蛋白调控Hippo/TAZ信号通路,进而促进骨髓间充质干细胞的成骨分化。CHEN等[28]在卵巢切除大鼠模型中发现,淫羊藿苷增加大鼠的骨量并降低了骨髓脂肪含量,促进成骨分化并抑制骨髓间充质干细胞的成脂分化,并且淫羊藿苷也能通过减弱雌激素受体α的作用来降低骨硬化蛋白表达,表明淫羊藿苷通过增强骨硬化蛋白甲基化来改善骨量并降低骨髓脂肪含量。金东升等[29]在兔骨髓间充质干细胞模型中证实,载淫羊藿苷缓释微球的三维支架能促进骨髓间充质干细胞的增殖,增加细胞活性增加,以10 μmol/L淫羊藿苷组效果最佳;淫羊藿苷促进了Ⅰ型胶原、骨钙素、Runt相关转录因子2、血管内皮生长因子蛋白表达。蔡叶等[30]在经D-半乳糖诱导的衰老骨髓间充质干细胞模型中发现,淫羊藿苷增加了衰老骨髓间充质干细胞上清液中碱性磷酸酶含量,并降低了P53的表达水平,可能通过调节成骨相关信号通路中转录因子(Foxo3a)/β-catenin的表达改善衰老骨髓间充质干细胞的成骨分化能力。张锦明等[31]研究发现,在经过连续皮下注射地塞米松5周的小鼠骨质疏松模型中,淫羊藿苷促进小鼠骨髓间充质干细胞的成骨分化、抑制小鼠骨髓间充质干细胞的成脂分化,改善骨髓微环境,促进骨小梁形成,缓解骨质疏松症。TENG等[32]实验发现,miR-335-5p在骨质疏松症骨组织中低表达,淫羊藿苷处理逆转了miR-335-5p抑制剂对骨髓间充质干细胞基质矿化的抑制作用,此外,肿瘤抑制基因蛋白被证实是miR-335-5p的下游效应子,在淫羊藿苷诱导过程中,肿瘤抑制基因蛋白的过表达逆转了miR-335-5p模拟物对骨髓间充质干细胞成骨分化的促进作用。 淫羊藿苷的成骨诱导作用主要通过调节骨髓间充质干细胞的定向分化实现的,淫羊藿苷可显著上调骨髓间充质干细胞中成骨关键因子Runt相关转录因子2、血管内皮生长因子及碱性磷酸酶的表达水平。进一步机制解析表明,淫羊藿苷的细胞分化调控效应依赖多条信号通路的协同作用,但存在浓度依赖性特征:当淫羊藿苷的浓度超过特定阈值时,成骨诱导作用显著减弱,而适宜浓度范围是发挥最佳骨形成效应的关键决定因素,这种双向调节特性最终指向优化骨髓间充质干细胞成骨分化过程并提升骨密度的核心目标,为后续实验设计提供了重要的剂量效应参考依据。淫羊藿苷调控骨髓间充质干细胞增殖、分化的作用机制详见表1。 2.2.2 淫羊藿苷对成骨细胞的影响 骨骼内骨组织的稳态依赖于新骨形成与陈旧骨吸收之间的精密调控,这种动态平衡机制确保了骨组织的持续更新及骨骼生物力学性能的维持[33]。当这种平衡被打破时,骨代谢失衡随即发生,进而引发骨质疏松症[34]。淫羊藿苷可显著促进成骨细胞功能活化,该作用机制主要体现在对骨髓间充质干细胞成骨定向分化的诱导效应。淫羊藿苷能加速成骨前体细胞增殖周期进程,并刺激骨钙素等骨基质蛋白合成,最终通过优化骨形成与骨吸收的偶联关系,实现骨量积累与骨质量改善的治疗目标。 ZHANG等[35]研究发现,在经地塞米松刺激的小鼠成骨细胞中,淫羊藿苷通过上调SPI1增加SMAD5表达,并能缓解SPI1缺失对细胞自噬与成骨分化的抑制作用。ZHU等[36]实验发现,在骨质疏松大鼠模型中,锶-淫羊藿苷复合涂层植入体周围可见更显著的新骨形成,表明锶-淫羊藿苷复合涂层能有效增强钛植入体的骨整合作用。ZHOU等[37]研究发现,在前成骨细胞MC3T3-E1模型中,淫羊藿苷能快速诱导胰岛素样生长因子1受体与雌激素受体α的相互作用,激活胰岛素样生长因子1受体信号通路及其下游蛋白激酶B磷酸化,淫羊藿苷对MC3T3-E1细胞的这种促进作用及其对成熟成骨细胞碱性磷酸酶活性的成骨效应均呈现胰岛素样生长因子1受体依赖性。陈绎志等[38]在兔股骨缺损动物模型中发现,淫羊藿苷可促进兔成骨细胞增殖,提高成骨细胞活性,增加骨形成。刘婷等[39]研究发现,淫羊藿苷能明显促进hFOB1.19人成骨细胞的增殖并促进细胞分泌碱性磷酸酶,提示淫羊藿苷对成骨细胞活性的促进作用可能是其防治骨质疏松症的途径之一。 综合现有研究,淫羊藿苷通过促进成骨细胞增殖与分化活性发挥抗骨质疏松症作用。研究进一步揭示,淫羊藿苷可通过上调转录因子SPI1基因表达增强受体激活蛋白SMAD5蛋白水平,逆转骨桥蛋白、骨钙素及Runt相关转录因子2等成骨标志物的表达抑制,从而同时实现减弱细胞自噬水平与缓解成骨分化障碍的双重调控效应。值得注意的是,基于骨组织工程技术的进展,锶元素与淫羊藿苷的协同应用在骨质疏松防治领域展现出显著的应用前景,为临床转化研究奠定了重要基础。淫羊藿苷调控成骨细胞增殖、分化的作用机制详见表2。 2.2.3 淫羊藿苷对破骨细胞的影响 破骨细胞作为骨吸收过程的核心效应细胞,属于多核巨型终末分化细胞,直接参与骨吸收,是骨组织吸收的主要功能细胞,与成骨细胞、骨细胞等形成动态调控体系,在细胞因子、生长因子及激素等多重调控信号的协同作用下,借助精密的分子信号传导通路实现骨吸收与骨形成的耦合平衡[40-41]。 SI等[42]研究发现,在经核因子κB受体活化因子配体诱导的RAW264.7细胞中,淫羊藿苷抑制E3泛素连接酶表达,并可通过减少内源性核转录因子E2相关因子的泛素化来延迟核转录因子E2相关因子降解,并且这些作用可被E3泛素连接酶过表达显著逆转,表明淫羊藿苷通过靶向E3泛素连接酶/核转录因子E2相关因子信号通路抑制破骨细胞生成,从而缓解骨质疏松症。CHENG等[43]实验结果表明,在经硫代乙酰胺诱导的破骨细胞中,淫羊藿苷抑制核因子κΒ配体受体激活剂、核因子κB受体激活剂、蛋白激酶(P38)、细胞外信号调节激酶、原癌基因c-Fos和活化T细胞核因子1的蛋白表达,结果表明淫羊藿苷可能通过下调蛋白激酶/细胞外信号调节激酶信号通路来抑制破骨细胞分化,阻止硫代乙酰胺诱导的骨质流失。朱在师等[44]研究发现,在可溶性核因子κB受体活化因子配体诱导的RAW264.7前破骨细胞模型中,知母皂苷Ⅱ及淫羊藿苷单独及联合应用均能够抑制破骨细胞分化,而不影响破骨细胞对成骨细胞的耦联作用,并且知母皂苷Ⅱ+淫羊藿苷联合应用能增强破骨细胞-成骨细胞偶联作用,该作用机制可能与促进破骨细胞源性偶联因子的表达有关。贺龙刚等[45]在体外RAW264.7细胞诱导破骨细胞的模型中发现,淫羊藿中的主要有效成分淫羊藿次苷Ⅰ及其代谢产物淫羊藿次苷Ⅱ在体外能通过抑制二聚体转录因子AP-1/活化T细胞核转录因子c1信号通路相关基因表达,提示可能通过抑制c-Fos蛋白调控靶基因转录因子Fra-1、转录因子Fra-2和活化T细胞核转录因子c1表达,进而抑制破骨细胞生成。 现有证据表明,破骨细胞功能异常是导致骨代谢失衡的核心病理环节,其中核因子κB受体活化因子配体/核因子κB受体活化因子信号轴作为调控破骨细胞活性的关键通路,已成为干预骨吸收过程的重要靶点。淫羊藿苷可通过抑制性调节核因子κB受体活化因子配体信号通路下调活化T细胞核转录因子c1、c-Fos、肿瘤坏死因子受体相关因子6及转化生长因子β等促破骨生成因子的表达水平,从而阻断破骨细胞前体的增殖分化进程。尽管现有实验数据已初步揭示淫羊藿苷与“破骨细胞-成骨细胞偶联”机制的潜在关联,但在成骨细胞信号网络中多蛋白因子的交互作用需进一步深入探讨。淫羊藿苷调控破骨细胞增殖、分化的作用机制详见表3。 2.3 淫羊藿苷防治骨质疏松症的相关信号通路研究 在淫羊藿苷防治骨质疏松症的研究中,Wnt/β-catenin、Notch、磷脂酰肌醇3-激酶/蛋白激酶B、丝裂原活化蛋白激酶及核因子κB受体活化因子/核因子κB受体活化因子配体/骨保护素通路为研究的重点,这些通路分别参与成骨细胞分化、破骨细胞活化及炎症反应的核心调控,是骨重建失衡的关键节点。淫羊藿苷通过激活Wnt/β-catenin促进成骨,抑制核因子κB受体活化因子配体/骨保护素失衡以减少破骨,同时调控Notch、磷脂酰肌醇3-激酶/蛋白激酶B及丝裂原活化蛋白激酶通路协"
"
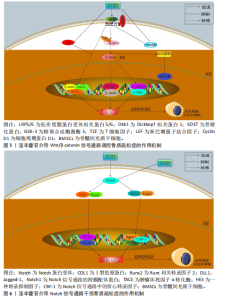
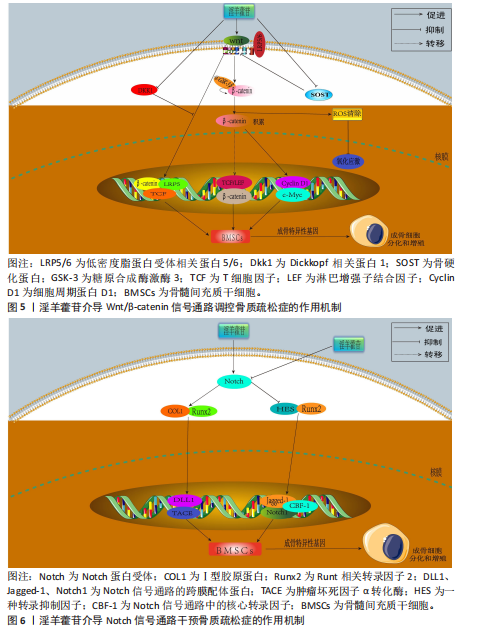
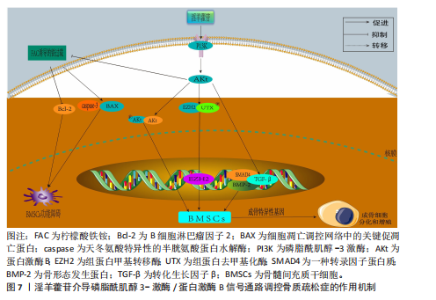
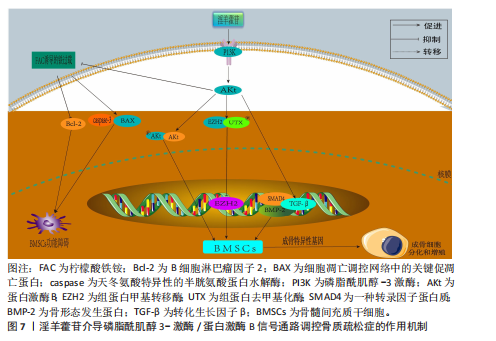
同平衡骨代谢。相关研究已明确淫羊藿苷对这些通路的干预作用,并且这些通路间存在交叉调控网络,能全面阐释淫羊藿苷多靶点作用机制;此外,这些通路是现有骨质疏松药物研发的重要靶点,为淫羊藿苷的临床转化提供理论依据。 2.3.1 Wnt/β-catenin 信号通路 Wnt-β-catenin信号轴在骨骼发育过程中发挥核心调控作用,它通过级联反应调控多个骨骼代谢及组织重塑相关关键因子的表达。Wnt-β-catenin信号传导过程始于Wnt蛋白与细胞膜表面Frizzled受体家族及低密度脂蛋白受体相关蛋白5/6共受体的特异性结合,这种相互作用主要发生在受体富含半胱氨酸的结构域,配体-受体复合物形成后触发细胞质内无规则蛋白的构象改变及功能活化。在Wnt信号通路激活状态下,蓬乱蛋白通过特异性蛋白互作界面拮抗轴抑制蛋白的分子支架功能,进而干扰由腺瘤性息肉病蛋白、轴抑制蛋白、糖原合成酶激酶3β及酪蛋白激酶1α构成的β-catenin降解复合体组 装[46-47]。在这一过程中,直接阻断β-catenin的磷酸化修饰及其后续泛素-蛋白酶体降解途径,导致该关键信号分子在胞质内异常蓄积,积累的β-catenin随即发生核转位,与T细胞因子/淋巴增强因子转录复合物形成功能性转录调控单元,通过启动下游靶基因的转录激活程序,最终实现其对细胞增殖、分化的调控作用[48-49]。 GAO等[50]在12周龄雌性SD大鼠骨髓间充质干细胞中发现,硬化蛋白基因诱导可显著抑制骨髓间充质干细胞的增殖和成骨分化,经淫羊藿苷处理后不仅增加了骨髓间充质干细胞的数量,还增强了成骨诱导过程中碱性磷酸酶活性和钙结节的形成;硬化蛋白下调了包括Runt相关转录因子2、β-catenin和原癌基因c-Myc在内的成骨基因以及抗氧化因子(Prdx1、转录因子Cata和Nqo1)的表达,经淫羊藿苷处理后上述指标表达得以恢复;淫羊藿苷通过激活Wnt/β-catenin通路来促进骨髓间充质干细胞的原位增殖和成骨分化。ZHANG等[51]在大鼠骨髓间充质干细胞模型中发现,不同浓度淫羊藿苷处理增加了碱性磷酸酶活性,并以剂量依赖性方式提高核心结合因子α1、骨形态发生蛋白2和骨形态发生蛋白4表达;此外,淫羊藿苷增加血清中了骨钙素、骨特异性碱性磷酸酶、Ⅰ型胶原蛋白N端肽、Ⅰ型胶原蛋白C端肽和抗酒石酸酸性磷酸酶5b水平,促进骨折部位骨钙素分泌,加速大鼠骨折愈合,该作用机制与激活Wnt1/β-catenin成骨信号通路相关。另有研究发现,淫羊藿苷通过激活Wnt/β-catenin信号通路调节去卵巢大鼠骨髓基质细胞成骨分化与脂肪分化的平衡,这可能是淫羊藿苷治疗绝经后骨质疏松症的重要分子机制[52]。 杨志烈等[53]研究发现,在环磷酰胺诱导的骨髓间充质干细胞成骨分化障碍模型中,经一定浓度的淫羊藿苷处理后细胞内Wnt4、β-catenin及细胞周期蛋白D1的mRNA水平显著升高,同时伴随β-catenin、细胞周期蛋白D1与c-Myc蛋白表达增强,而Dickkopf 相关蛋白1表达呈现下调趋势,该作用可能通过激活Wnt/β-catenin信号通路实现的。曾建春等[54]在人骨髓间充质干细胞模型中发现,含40 μg/L淫羊藿苷诱导液诱导细胞向成骨细胞分化,并且显著提高Wnt3a、β-catenin蛋白表达,该作用机制是通过激活Wnt/β-catenin信号通路实现的。ZENG等[55]在乙醇干预下的MC3T3-E1细胞模型中发现,淫羊藿苷增加了细胞内相对碱性磷酸酶面积,上调被酒精抑制的成骨相关蛋白和mRNAs表达水平,促进成骨分化;此外淫羊藿苷还降低了酒精升高的活性氧水平,减少了酒精干预诱导的成骨细胞凋亡,该作用机制是通过调节Wnt/β-catenin信号通路实现的。淫羊藿苷介导 Wnt/β-catenin信号通路调控骨质疏松症的作用机制,详见图5。 2.3.2 Notch信号通路 Notch信号通路在细胞增殖、分化及凋亡的调控网络中发挥关键作用,尤其在成骨分化早期阶段呈现促分化效应,而分化后期则表现为对成骨细胞增殖的抑制作用。当骨髓间充质干细胞的Notch通路被激活时,该通路通过维持干细胞未分化状态来促进细胞增殖,而阻断骨髓间充质干细胞内的Notch信号传导可诱导干细胞向成骨或软骨细胞定向分化[56]。Notch信号通路配体Jagged-1的过表达可通过延缓细胞衰老进程加速骨折早期修复,并显著提升成骨前体细胞的分化潜能[57]。通过抑制Notch信号通路活性可增强破骨细胞增殖活性并减少细胞凋亡;而Notch-1受体过度活化通过上调δ样配体3及Jagged-1表达水平发挥对破骨细胞的抑制效应[58-60]。 BIAN等[61]在去卵巢术诱导大鼠骨质疏松模型中发现,淫羊藿苷可通过调控Delta样配体1及肿瘤坏死因子α转化酶的表达水平激活Notch信号传导通路,该过程进一步诱导骨钙素、Ⅰ型胶原蛋白及Runt相关转录因子2分泌量增加,促进骨髓间充质干细胞向成骨分化。邓宇等[62]在大鼠骨髓间充质干细胞模型中发现,淫羊藿苷可显著上调HES、Runt相关转录因2 mRNA表达,同时促进Jagged-1、Notch-1、CBF1蛋白表达水平,进一步激活Notch信号通路,促进骨髓间充质干细胞向成骨细胞分化,进而改善骨质疏松症。徐娅等[63]在大鼠骨髓间充质干细胞的体外模型中发现,淫羊藿苷能够促进骨髓间充质干细胞向成骨分化,通过下调 Notch信号核心结合蛋白CBF1与通路中关键蛋白Notch-1的表达抑制Notch信号通路,从而发挥抗骨质疏松症作用。周亚妮等[64]在去卵巢骨质疏松症大鼠骨髓间充质干细胞模型中发现,淫羊藿苷通过上调碱性磷酸酶和骨钙素水平及抑制Notch通路中Notch-1、CBF1、Jagged-1蛋白表达,促进骨髓间充质干细胞的增殖和成骨细胞分化,从而达到防治骨质疏松症作用。淫羊藿苷介导Notch 信号通路干预骨质疏松症的作用机制,详见图6。 2.3.3 磷脂酰肌醇3-激酶/蛋白激酶B信号通路 磷脂酰肌醇3-激酶/蛋白激酶B信号传导系统普遍存在于各类细胞中,作为细胞内外信号整合枢纽,通过多层级调控网络参与细胞生长调控、增殖维持、分化定向及细胞骨架动态重排等关键生物学过程[65]。蛋白激酶B蛋白家族包含蛋白激酶B1、蛋白激酶B2和蛋白激酶B3三种亚型。作为磷脂酰肌醇信号通路的关键效应分子,磷脂酰肌醇3-激酶通过与蛋白激酶B的N端PH结构域结合诱导其构象重塑,使Thr308和Ser473两个关键磷酸化位点暴露并发生双重磷酸化修饰,这种共价修饰是蛋白激酶B活性激活的分子基础[66]。激活态蛋白激酶B通过磷酸化级联反应调控下游效应蛋白,包括哺乳动物雷帕霉素靶蛋白复合物1、MDM2原癌蛋白、Bcl-2相关死亡促进因子、核因子κB、糖原合成酶激酶3β、半胱氨酸天冬氨酸蛋白酶9、核糖体蛋白S6激酶及FoxO转录因子家族等,进而促进细胞新陈代谢、生长、增殖、凋亡和血管生成等[67-68]。 YAO等[69]在经柠檬酸铁铵诱导的铁过载骨髓间充质干细胞模型中发现,caspase-3和BAX蛋白表达升高,Bcl-2蛋白表达降低,淫羊藿苷处理可降低caspase-3和BAX蛋白表达、升高Bcl-2蛋白表达,表明淫羊藿苷能够防止铁过载诱导的骨髓间充质干细胞功能障碍,该作用机制与淫羊藿苷减弱铁过载诱导磷脂酰肌醇3-激酶/蛋白激酶B通路失活相关。JI等[70]在经地塞米松诱导的骨髓间充质干细胞模型中发现,淫羊藿苷通过激活磷脂酰肌醇3-激酶/蛋白激酶B信号传导上调组蛋白去甲基化酶并抑制组蛋白甲基转移酶2表达,促进地塞米松处理骨髓间充质干细胞的成骨分化。范凯健等[71]在MC3T3-E1细胞模型中发现,淫羊藿苷干预可以促进钙沉积、提高碱性磷酸酶活性,提高SMAD4、骨形态发生蛋白2、转化生长因子β蛋白表达,明显促进细胞的增殖与迁移,作用机制可能是淫羊藿苷通过促进磷脂酰肌醇3-激酶/蛋白激酶B信号通路的磷酸化有关。郭晓宇等[72]在大鼠骨髓间充质干细胞模型中发现,淫羊藿苷处理24 h后细胞内碱性磷酸酶基因表达水平、蛋白激酶B磷酸化表达水平升高,表明淫羊藿苷通过激活磷脂酰肌醇3-激酶/蛋白激酶B途径来促进大鼠骨髓间充质干细胞的成骨分化。淫羊藿苷介导磷脂酰肌醇3-激酶/蛋白激酶B信号通路调控骨质疏松症的作用机制,详见图7。 2.3.4 丝裂原活化蛋白激酶信号通路 丝裂原活化蛋白激酶信号通路作为细胞生理学调控及免疫应答的核心信号模块,是真核生物信号转导系统的关键结构单元,该通路通过级联反应介导并调控细胞分化、增殖、细胞凋亡、炎症反应和氧化应激等[73]。现有研究表明,丝裂原活化蛋白激酶信号通路在骨代谢调控网络中发挥核心调控作用,该通路通过精确调控破骨细胞生成关键下游效应分子c-Fos和活化T细胞核转录因子c1的表达活性形成级联调控效应,最终驱动破骨细胞分化进程[74-75]。丝裂原活化蛋白激酶信号转导系统由细胞外信号调节激酶、c-Jun末端激酶和P38丝裂原活化蛋白激酶等平行信号轴构成,其磷酸化参与调控核因子κB受体活化因子配体诱导的破骨细胞生成及骨吸收过程,而丝裂原活化蛋白激酶特异性抑制剂可有效阻断核因子κB受体活化因子配体介导的破骨细胞分化效应[76]。 HSIEH等[77]在脂多糖诱导的破骨细胞模型中发现,淫羊藿苷抑制脂多糖诱导的骨吸收和白细胞介素6和肿瘤坏死因子α表达,并且抑制脂多糖介导的P38和c-Jun末端激酶激活,表明淫羊藿苷通过丝裂原活化蛋白激酶通路抑制破骨细胞分化,防止炎症性骨质流失。DENG等[78]在苯和环磷酰胺诱导再生障碍性贫血大鼠骨髓间充质干细胞模型中发现,淫羊藿苷以剂量依赖性方式促进骨髓间充质干细胞的增殖,下调骨髓间充质干细胞中磷酸化P38/P38、磷酸化c-Jun末端激酶/c-Jun末端激酶和磷酸化细胞外信号调节激酶/细胞外信号调节激酶的蛋白水平,抑制骨髓间充质干细胞的脂肪分化,表明淫羊藿苷通过抑制丝裂原活化蛋白激酶通路促进骨髓间充质干细胞的增殖并抑制细胞凋亡和脂肪生成。QIN等[79]在大鼠骨髓间充质干细胞模型中发现,不同浓度淫羊藿苷处理可显著提高骨髓间充质干细胞中细胞外信号调节激酶和P38激酶的磷酸化,并且上调丝裂原活化蛋白激酶下游转录因子Elk1和骨髓细胞瘤病毒癌基因c-Myc的表达,并且最佳处理质量浓度度为 320 μg/L,表明淫羊藿苷通过激活细胞外信号调节激酶和P38丝裂原活化蛋白激酶信号传导促进大鼠骨髓间充质干细胞的增殖。XU等[80]在核因子κB受体活化因子配体诱导RAW264.7细胞向破骨细胞分化的实验中发现,经用不同浓度(0,10,10和10 mol/L)淫羊藿苷处理后,淫羊藿苷以剂量依赖性方式显著抑制了核因子κB受体活化因子配体诱导的活化T细胞核转录因子c1、c-Fos、抗酒石酸酸性磷酸酶和组织蛋白酶K的表达,表明淫羊藿苷在体外抑制核因子κB受体活化因子配体诱"
"
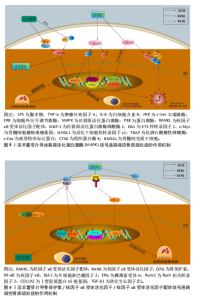
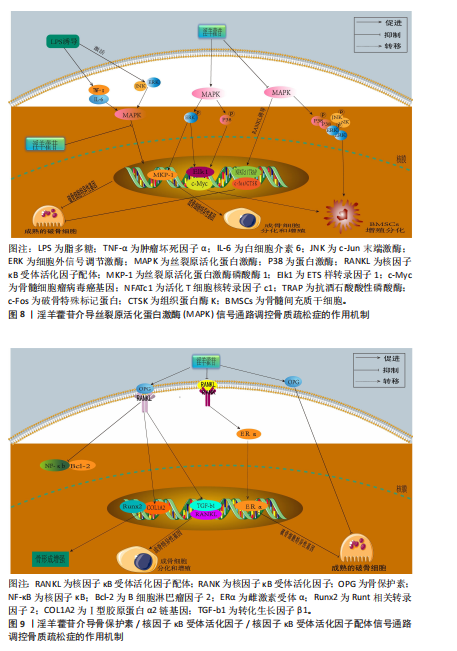
导的破骨细胞形成和功能,该作用机制是通过抑制丝裂原活化蛋白激酶信号通路的激活来实现的。刘尚全等[81]在经地塞米松诱导的成骨细胞模型中发现,地塞米松促进丝裂原活化蛋白激酶磷酸酶1的表达、抑制丝裂原活化蛋白激酶的磷酸化、减弱丝裂原活化蛋白激酶信号途径的活性,从而抑制了成骨细胞的增殖和分化;淫羊藿苷处理使细胞外信号调节激酶磷酸化增加,同时抑制丝裂原活化蛋白激酶磷酸酶1的表达,表明淫羊藿苷通过丝裂原活化蛋白激酶信号途径促进了细胞的增殖分化,逆转了地塞米松对成骨细胞增殖分化的抑制作用。淫羊藿苷介导丝裂原活化蛋白激酶信号通路调控骨质疏松症的作用机制,详见图8。 2.3.5 骨保护素/核因子κB受体活化因子/核因子κB受体活化因子配体通路 骨保护素作为成骨细胞及骨髓间充质干细胞分泌的关键骨形成调控蛋白,通过拮抗核因子κB受体活化因子配体生物学活性参与破骨细胞分化成熟调控及骨代谢稳态维持。骨保护素/核因子κB受体活化因子配体/核因子κB受体活化因子信号轴构成破骨细胞生成及骨重塑过程的核心调控通路,在破骨细胞功能活化与骨组织动态平衡中发挥枢纽作用[82]。作为骨保护素/核因子κB受体活化因子配体/核因子κB受体活化因子信号轴的核心调控节点,核因子κB受体活化因子配体与其膜受体核因子κB受体活化因子的特异性结合可激活破骨细胞分化成熟程序,通过维持破骨细胞存活状态维持骨吸收活性。在该通路调控网络中,骨保护素作为可溶性诱骗受体,通过竞争性结合核因子κB受体活化因子配体分子有效阻断核因子κB受体活化因子配体-核因子κB受体活化因子信号转导,从而抑制破骨细胞功能活性并诱导其发生程序性死亡,最终实现骨吸收过程的负向调控[83]。因此,骨保护素/核因子κB受体活化因子配体比值是破骨细胞分化、成熟的重要指标,也是保持骨吸收和骨形成平衡状态的关键[84-85]。 ZHANG等[86]在双侧卵巢切除诱导的绝经后骨质疏松性骨折大鼠中发现,淫羊藿苷通过骨保护素/核因子κB受体活化因子配体通路促进Runt相关转录因子2和Ⅰ型胶原蛋白α2链基因的表达水平,抑制血清骨翻转生物标志物的表达水平,表明淫羊藿苷可通过骨保护素/ 核因子κB受体活化因子配体信号传导促进去卵巢大鼠骨折愈合。ZHANG等[87]在小鼠成骨细胞MC3T3-E和小鼠破骨细胞RAW264.7共培养系统中发现,一定浓度的淫羊藿苷处理可提高骨保护素、核因子κB受体活化因子配体基因和蛋白的表达,降低核因子κB基因和蛋白的表达,增加碱性磷酸酶、转化生长因子β1和核因子κB受体活化因子配体基因的表达,降低核因子κB受体活化因子基因的表达,表明淫羊藿苷促进MC3T3-E1细胞的成骨活性、抑制RAW264.7 细胞的破骨细胞活性。李伟娟等[88]在核因子κB受体活化因子配体诱导小鼠RAW264.7细胞破骨分化的模型中发现,淫羊藿苷显著抑制核因子κB受体活化因子配体诱导RAW264.7细胞分化成破骨细胞,这种抑制作用可通过升高雌激素受体α表达而直接或间接降低核因子κB受体活化因子表达,进而降低核因子κB受体活化因子配体/核因子κB受体活化因子信号水平,最终降低破骨细胞形成数量和破骨细胞骨吸收活性。吴祖锋等[89]在大鼠双侧卵巢切除构建的骨质疏松症模型中发现,低、中、高剂量(200,400,800 mg/kg)淫羊藿苷灌胃干预可显著提升大鼠骨保护素蛋白表达水平、上调核因子κB受体活化因子配体/核因子κB受体活化因子信号通路活性,进而抑制破骨细胞分化,发挥防治骨质疏松作用。马小妮等[90]在大鼠骨髓间充质干细胞模型中发现,10-5 mol/L淫羊藿苷干预可显著提升骨保护素蛋白表达水平、骨保护素/核因子κB受体活化因子配体比值,通过阻断破骨细胞前体分化及成熟破骨细胞功能活性发挥抗骨吸收效应,表明淫羊藿苷通过调控骨保护素/核因子κB受体活化因子配体信号通路影响骨吸收,从而发挥抗骨质疏松症的作用。淫羊藿苷介导骨保护素/核因子κB受体活化因子/核因子κB受体活化因子配体信号通路调控骨质疏松症的作用机制,详见图9。"
| [1] ZOU Z, LIU W, CAO L, et al. Advances in the occurrence and biotherapy of osteoporosis. Biochem Soc Trans. 2020;48(4):1623-1636. [2] ELONHEIMO H, LANGE R, TOLONEN H, et al. Environmental Substances Associated with Osteoporosis-A Scoping Review. Int J Environ Res Public Health. 2021;18(2):738. [3] 中国骨质疏松症流行病学调查及“健康骨骼”专项行动结果发布[J].中华骨质疏松和骨矿盐疾病杂志,2019,12(4):317-318. [4] 中华医学会骨质疏松和骨矿盐疾病分会,章振林.原发性骨质疏松症诊疗指南(2022)[J].中国全科医学,2023,26(14):1671-1691. [5] JOHNSTON CB, DAGAR M. Osteoporosis in Older Adults. Med Clin North Am. 2020;104(5): 873-884. [6] SLEEMAN A, CLEMENTS JN. Abaloparatide: A new pharmacological option for osteoporosis. Am J Health Syst Pharm. 2019;76(3):130-135. [7] MEGAPANOU E, FLORENTIN M, MILIONIS H, et al. Drug-Induced Hypophosphatemia: Current Insights. Drug Saf. 2020;43(3):197-210. [8] SUN P, ZHANG C, HUANG Y, et al. Jiangu granule ameliorated OVX rats bone loss by modulating gut microbiota-SCFAs-Treg/Th17 axis. Biomed Pharmacother. 2022;150:112975. [9] WANG Z, WANG D, YANG D, et al. The effect of icariin on bone metabolism and its potential clinical application. Osteoporos Int. 2018;29(3):535-544. [10] QIAN HQ, WU DC, LI CY, et al. A systematic review of traditional uses, phytochemistry, pharmacology and toxicity of Epimedium koreanum Nakai. J Ethnopharmacol. 2024; 318(Pt B):116957. [11] ZHAO M, XU P, SHI W, et al. Icariin exerts anti-tumor activity by inducing autophagy via AMPK/mTOR/ULK1 pathway in triple-negative breast cancer. Cancer Cell Int. 2024;24(1):74. [12] LEI X, WEN D, HUANG Z, et al. Icariin attenuates oxidative stress via SIRT1/PGC-1α pathway in SAH mice. Exp Neurol. 2025;390:115303. [13] ZHANG X, HAN N, LI G, et al. Local icariin application enhanced periodontal tissue regeneration and relieved local inflammation in a minipig model of periodontitis. Int J Oral Sci. 2018;10(2):19. [14] GAO L, ZHANG SQ. Antiosteoporosis Effects, Pharmacokinetics, and Drug Delivery Systems of Icaritin: Advances and Prospects. Pharmaceuticals (Basel). 2022;15(4):397. [15] SHA M, CAO A, YANG S, et al. [Determination of icariin in Epimedium koreanum nakai by high performance liquid chromatography]. Se Pu. 1997;15(2):166-177. [16] XIAO Q, CHEN A, GUO F. Effects of Icariin on expression of OPN mRNA and type I collagen in rat osteoblasts in vitro. J Huazhong Univ Sci Technolog Med Sci. 2005;25(6):690-702. [17] CHEN KM, GE BF, LIU XY, et al. Icariin inhibits the osteoclast formation induced by RANKL and macrophage-colony stimulating factor in mouse bone marrow culture. Pharmazie. 2007;62(5):388-891. [18] MOK SK, CHEN WF, LAI WP, et al. Icariin protects against bone loss induced by oestrogen deficiency and activates oestrogen receptor-dependent osteoblastic functions in UMR 106 cells. Br J Pharmacol. 2010;159(4):939-949. [19] LIANG W, LIN M, LI X, et al. Icariin promotes bone formation via the BMP-2/Smad4 signal transduction pathway in the hFOB 1.19 human osteoblastic cell line. Int J Mol Med. 2012;30(4):889-895. [20] ZHANG D, FONG C, JIA Z, et al. Icariin Stimulates Differentiation and Suppresses Adipocytic Transdifferentiation of Primary Osteoblasts Through Estrogen Receptor-Mediated Pathway. Calcif Tissue Int. 2016;99(2):187-198. [21] LIANG X, HOU Z, XIE Y, et al. Icariin promotes osteogenic differentiation of bone marrow stromal cells and prevents bone loss in OVX mice via activating autophagy. J Cell Biochem. 2019;120(8):13121-13132. [22] BALLHAUSE TM, JIANG S, XIE W, et al. Fracture healing in a mouse model of Hajdu-Cheney-Syndrome with high turnover osteopenia results in decreased biomechanical stability. Sci Rep. 2023;13(1):11418. [23] SANTOS TA, RIBEIRO JL, BATTISTELLI LS, et al. Limosilactobacillus reuteri with menaquinone-7 improves bone biomechanics and microarchitecture in ovariectomized mice: preliminary study. J Bone Miner Metab. 2025;43(4):335-347. [24] WANIONOK NE, COLAREDA GA, FERNANDEZ JM. Humulus lupulus Promoting Osteoblast Activity and Bone Integrity: Effects and Mechanisms. Biology (Basel). 2025;14(5):582. [25] CLAVIN NW, FERNANDEZ J, SCHÖNMEYR BH, et al. Fractionated doses of ionizing radiation confer protection to mesenchymal stem cell pluripotency. Plast Reconstr Surg. 2008;122(3):739-748. [26] BUDGUDE P, KALE V, VAIDYA A. Microvesicles and exosomes isolated from murine bone marrow-derived mesenchymal stromal cells primed with p38MAPK inhibitor differentially regulate hematopoietic stem cell function. Artif Cells Nanomed Biotechnol. 2025;53(1):122-137. [27] HAN YD, ZHANG HF, XU YT, et al.[Mechanism of icariin in promoting osteogenic differentiation of BMSCs and improving bone metabolism disorders through caveolin-1/Hippo signaling pathway]. Zhongguo Zhong Yao Za Zhi. 2025; 50(3):600-608. [28] CHEN X, LIU X, WAN J, et al. Icariin Facilitates Osteogenic Differentiation and Suppresses Adipogenic Differentiation of Bone Marrow Mesenchymal Stem Cells by Enhancing SOST Methylation in Postmenopausal Osteoporosis. J Gene Med. 2025;27(1):e70010. [29] 金东升,赵张红,朱子银,等.淫羊藿苷缓释微球三维支架对兔骨髓间充质干细胞成骨分化的影响[J].中国组织工程研究,2026, 30(7):1658-1668. [30] 蔡叶,王明飞,张磊,等.淫羊藿苷对衰老骨髓间充质干细胞成骨的影响[J].中国骨质疏松杂志,2023,29(2):162-167. [31] 张锦明,田滢舟,赵玲,等.淫羊藿苷促进骨髓间充质干细胞成骨分化缓解小鼠骨质疏松的机制[J].中国组织工程研究,2022, 26(19):2991-2996. [32] TENG JW, BIAN SS, KONG P, et al. Icariin triggers osteogenic differentiation of bone marrow stem cells by up-regulating miR-335-5p. Exp Cell Res. 2022;414(2):113085. [33] 王想福,孙凤歧,叶丙霖,等.破骨细胞与骨质疏松症的关系研究进展[J].中国骨质疏松杂志,2015,21(11):1420-1424. [34] CHU W, PENG W, LU Y, et al. PRMT6 Epigenetically Drives Metabolic Switch from Fatty Acid Oxidation toward Glycolysis and Promotes Osteoclast Differentiation During Osteoporosis. Adv Sci (Weinh). 2024; 11(40):e2403177. [35] ZHANG J, MAO Y, RAO J. The SPI1/SMAD5 cascade in the promoting effect of icariin on osteogenic differentiation of MC3T3-E1 cells: a mechanism study. J Orthop Surg Res. 2024;19(1):444. [36] ZHU Y, ZHENG T, WEN LM, et al. Osteogenic capability of strontium and icariin-loaded TiO2 nanotube coatings in vitro and in osteoporotic rats. J Biomater Appl. 2021;35(9): 1119-1131. [37] ZHOU L, POON CC, WONG KY, et al. Icariin ameliorates estrogen-deficiency induced bone loss by enhancing IGF-I signaling via its crosstalk with non-genomic ERα signaling. Phytomedicine. 2021;82:153413. [38] 陈绎志,董忠,杨俊,等.淫羊藿苷对兔成骨细胞的调控[J].中国老年学杂志,2023, 43(8):1937-1939. [39] 刘婷,曹春雨,郝然,等.黔岭淫羊藿总黄酮类成分对hFOB1.19人SV40转染成骨细胞活性的影响[J].中国实验方剂学杂志, 2013,19(2):267-270. [40] SONG L, ZHOU Y, QU L, et al. Exploring Effects and Mechanism of Ingredients of Herba Epimedii on Osteogenesis and Osteoclastogenesis In Vitro. Comb Chem High Throughput Screen. 2024;27(19): 2824-2837. [41] ZHANG D, ZHANG J, FONG C, et al. Herba epimedii flavonoids suppress osteoclastic differentiation and bone resorption by inducing G2/M arrest and apoptosis. Biochimie. 2012; 94(12):2514-2522. [42] SI Y, LI Y, GU K, et al. Icariin ameliorates osteoporosis in ovariectomized rats by targeting Cullin 3/Nrf2/OH pathway for osteoclast inhibition. Biomed Pharmacother. 2024;173:116422. [43] CHENG L, JIN X, SHEN H, et al. Icariin attenuates thioacetamideinduced bone loss via the RANKLp38/ERKNFAT signaling pathway. Mol Med Rep. 2022;25(4):126. [44] 朱在师,黄泽灵,蔡伟业,等.基于“阴阳平衡”探讨知母皂苷BⅡ-淫羊藿苷协同对破骨细胞-成骨细胞耦联功能的影响[J].中国实验方剂学杂志,2025,31(18):48-57. [45] 贺龙刚,高奥,邱煌沛,等.淫羊藿次苷Ⅰ及其代谢产物淫羊藿次苷Ⅱ通过AP-1/NFATc1信号通路调控破骨细胞生成[J].中华中医药杂志,2017,32(3):1299-1302. [46] ZHAO Y, LIU J, ZHANG Y, et al. Mir-381-3p aggravates ovariectomy-induced osteoporosis by inhibiting osteogenic differentiation through targeting KLF5/Wnt/β-catenin signaling pathway. J Orthop Surg Res. 2024; 19(1):480. [47] ZHENG S, HU G, ZHENG J, et al. Osthole accelerates osteoporotic fracture healing by inducing the osteogenesis-angiogenesis coupling of BMSCs via the Wnt/β-catenin pathway. Phytother Res. 2024;38(8):4022-4035. [48] PUJIA A, RUSSO C, MAUROTTI S, et al. Bergamot Polyphenol Fraction Exerts Effects on Bone Biology by Activating ERK 1/2 and Wnt/β-Catenin Pathway and Regulating Bone Biomarkers in Bone Cell Cultures. Nutrients. 2018;10(9):1305. [49] HONG G, HE X, SHEN Y, et al. Chrysosplenetin promotes osteoblastogenesis of bone marrow stromal cells via Wnt/β-catenin pathway and enhances osteogenesis in estrogen deficiency-induced bone loss. Stem Cell Res Ther. 2019; 10(1):277. [50] GAO J, XIANG S, WEI X, et al. Icariin Promotes the Osteogenesis of Bone Marrow Mesenchymal Stem Cells through Regulating Sclerostin and Activating the Wnt/β-Catenin Signaling Pathway. Biomed Res Int. 2021;2021:6666836. [51] ZHANG XY, CHEN YP, ZHANG C, et al. Icariin Accelerates Fracture Healing via Activation of the WNT1/β-catenin Osteogenic Signaling Pathway. Curr Pharm Biotechnol. 2020;21(15): 1645-1653. [52] XU YX, WU CL, WU Y, et al. Epimedium-derived flavonoids modulate the balance between osteogenic differentiation and adipogenic differentiation in bone marrow stromal cells of ovariectomized rats via Wnt/β-catenin signal pathway activation. Chin J Integr Med. 2012;18(12):909-917. [53] 杨志烈,王成龙,赵东峰,等.淫羊藿苷对环磷酰胺化疗导致小鼠骨髓间充质干细胞成骨分化障碍的保护作用[J].中国组织工程研究,2016,20(6):777-784. [54] 曾建春,曾意荣,樊粤光,等.淫羊藿甙诱导MSCs向成骨细胞分化过程中对Wnt信号通路的影响[J].广州中医药大学学报,2014, 31(4):607-611,678. [55] ZENG Q, CHEN YP, SONG SL, et al. [Icariin promotes alcohol-inhibited osteogenic differentiation of MC3T3-1-E1 cells by regulating LAP autophagy]. Zhongguo Zhong Yao Za Zhi. 2025;50(3):590-599. [56] ZANOTTI S, CANALIS E. Notch Signaling and the Skeleton. Endocr Rev. 2016;37(3): 223-253. [57] TIAN Y, XU Y, XUE T, et al. Notch activation enhances mesenchymal stem cell sheet osteogenic potential by inhibition of cellular senescence. Cell Death Dis. 2017; 8(2):e2595. [58] TAO ZS, LI TL, XU HG, et al. Hydrogel contained valproic acid accelerates bone-defect repair via activating Notch signaling pathway in ovariectomized rats. J Mater Sci Mater Med. 2021;33(1):4. [59] XU C, DINH VV, KRUSE K, et al. Induction of osteogenesis by bone-targeted Notch activation. Elife. 2022;11:e60183. [60] ZHU Z, WANG Z, MA C, et al. Isopsoralen promotes osteogenic differentiation of human jawbone marrow mesenchymal cells through Notch signaling pathway. Ann Anat. 2023;250:152156. [61] BIAN Q, HUANG JH, LIU SF, et al. Different molecular targets of Icariin on bMSCs in CORT and OVX-rats. Front Biosci (Elite Ed). 2012;4(4): 1224-1236. [62] 邓宇,陈廖斌.淫羊藿苷通过激活Notch信号通路促进骨髓间充质干细胞向成骨细胞分化的实验研究[J].中医学报,2017,32(12): 2393-2398,2403. [63] 徐娅,王攀攀,许青青,等. 淫羊藿苷在促大鼠骨髓间充质干细胞骨向分化过程中对Notch信号通路Notch1、CBF1蛋白表达的影响[C]//第十三次全国中西医结合虚证与老年医学学术研讨会论文集,2013:103-111. [64] 周亚妮,张晓文,王丹.淫羊藿苷抑制Notch信号通路促进骨髓间充质干细胞分化研究[J].商洛学院学报,2024,38(4):70-75. [65] CHANG L, ZHAO D, LIU HB, et al. Activation of sonic hedgehog signaling enhances cell migration and invasion by induction of matrix metalloproteinase-2 and -9 via the phosphoinositide-3 kinase/AKT signaling pathway in glioblastoma. Mol Med Rep. 2015; 12(5):6702-6710. [66] SOLIT DB, BASSO AD, OLSHEN AB, et al. Inhibition of heat shock protein 90 function down-regulates Akt kinase and sensitizes tumors to Taxol. Cancer Res. 2003;63(9): 2139-2144. [67] HUANG H, JI F, QI G, et al. Rehmannioside A promotes the osteoblastic differentiation of MC3T3-E1 cells via the PI3K/AKT signaling pathway and inhibits glucocorticoid-induced bone loss in vivo. J Pharmacol Sci. 2024; 156(4):247-257. [68] XU K, FEI W, GAO W, et al. SOD3 regulates FLT1 to affect bone metabolism by promoting osteogenesis and inhibiting adipogenesis through PI3K/AKT and MAPK pathways. Free Radic Biol Med. 2024;212:65-79. [69] YAO X, JING X, GUO J, et al. Icariin Protects Bone Marrow Mesenchymal Stem Cells Against Iron Overload Induced Dysfunction Through Mitochondrial Fusion and Fission, PI3K/AKT/mTOR and MAPK Pathways. Front Pharmacol. 2019;10:163. [70] JI W, GONG G, LIU Y, et al. Icariin promotes osteogenic differentiation of bone marrow mesenchymal stem cells (BMSCs) by activating PI3K-AKT-UTX/EZH2 signaling in steroid-induced femoral head osteonecrosis. J Orthop Surg Res. 2025;20(1):290. [71] 范凯健,钮艾雯,吴辉辉.淫羊藿苷通过PI3K/AKT信号通路促进MC3T3-E1细胞的增殖分化[J].中南药学,2025,23(2):417-422. [72] 郭晓宇,李唯,陈克明,等.淫羊藿苷通过PI3K/AKT-eNOS信号途径促进大鼠骨髓基质细胞的成骨性分化[J].中国药理学通报, 2013,29(7):966-970. [73] MENG X, ZHANG S. MAPK cascades in plant disease resistance signaling. Annu Rev Phytopathol. 2013;51:245-266. [74] HTIKE K, YOSHIDA K, EGUCHI T, et al. Herbal medicine Ninjinyoeito inhibits RANKL-induced osteoclast differentiation and bone resorption activity by regulating NF-κB and MAPK pathway. J Oral Biosci. 2024;66(4):49-57. [75] LIU C, ZUO M, ZHAO J, et al. DPHB inhibits osteoclastogenesis by suppressing NF-κB and MAPK signaling and alleviates inflammatory bone destruction. Int Immunopharmacol. 2025;152:114377. [76] LI X, GAO C, ZHOU K, et al. Dendrobine Ameliorates Glucocorticoid-Induced Osteoporosis by Promoting Osteogenesis through JNK/p38 MAPK Pathway Activation and GR Nuclear Translocation Inhibition. J Agric Food Chem. 2024;72(30):16739-16748. [77] HSIEH TP, SHEU SY, SUN JS, et al. Icariin inhibits osteoclast differentiation and bone resorption by suppression of MAPKs/NF-κB regulated HIF-1α and PGE(2) synthesis. Phytomedicine. 2011;18(2-3):176-185. [78] DENG S, ZENG Y, XIANG J, et al. Icariin protects bone marrow mesenchymal stem cells in aplastic anemia by targeting MAPK pathway. Mol Biol Rep. 2022;49(9):8317-8324. [79] QIN S , ZHOU W, LIU S, et al. Icariin stimulates the proliferation of rat bone mesenchymal stem cells via ERK and p38 MAPK signaling. Int J Clin Exp Med. 2015;8(5):7125-7133. [80] XU Q, CHEN G, LIU X, et al. Icariin inhibits RANKL-induced osteoclastogenesis via modulation of the NF-κB and MAPK signaling pathways. Biochem Biophys Res Commun. 2019;508(3):902-906. [81] 刘尚全,杨颖,周丽斌,等.淫羊藿甙逆转地塞米松抑制成骨细胞分化及其机制[J].中华内分泌代谢杂志,2006,22(3):218-221. [82] NI X, WU B, LI S, et al. Equol exerts a protective effect on postmenopausal osteoporosis by upregulating OPG/RANKL pathway. Phytomedicine. 2023;108: 154509. [83] 叶子丰,戎宽,张信成,等.基于OPG/RANKL/RANK信号通路探讨益肾健骨方防治绝经后骨质疏松的作用机制[J].中国骨质疏松杂志,2023,29(1):23-28,57. [84] 谢忠建.从OPG-RANKL-RANK通路到骨质疏松症分子靶向治疗[J].中华骨质疏松和骨矿盐疾病杂志,2022,15(2):126-134. [85] 高城翰,刘晓炜,关雪峰.补肾法对去卵巢大鼠OPG/RANK/RANKL信号通路影响的Meta分析[J].中国实验方剂学杂志,2021, 27(20):172-179. [86] ZHANG Y, HAN B, WEI Y, et al. Icariin Promotes Fracture Healing in Ovariectomized Rats. Med Sci Monit. 2020;26:e924554. [87] ZHANG S, FENG P, MO G, et al. Icariin influences adipogenic differentiation of stem cells affected by osteoblast-osteoclast co-culture and clinical research adipogenic. Biomed Pharmacother. 2017;88:436-442. [88] 李伟娟,谢保平,石丽颖,等.从ERα/RANK通路探讨淫羊藿苷抑制破骨细胞分化作用[J].中国实验方剂学杂志,2017, 23(7):121-126. [89] 吴祖锋,袁垒,吴风晴,等.淫羊藿苷对骨质疏松症模型大鼠OPG/RANKL/RANK轴系统影响的实验研究[J].甘肃中医药大学学报, 2016,33(3):4-7. [90] 马小妮,葛宝丰,陈克明,等.淫羊藿苷通过OPG/RANKL信号途径调节骨吸收的机理研究[J].中国骨质疏松杂志,2013,19(1):1-5. |
| [1] | Liu Wenlong, Dong Lei, Xiao Zhengzheng, Nie Yu. Finite element analysis of tibial prosthesis loosening after fixed-bearing unicompartmental knee arthroplasty for osteoporosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2191-2198. |
| [2] | Chen Long, Wang Xiaozhen, Xi Jintao, Lu Qilin. Biomechanical performance of short-segment screw fixation combined with expandable polyetheretherketone vertebral body replacement in osteoporotic vertebrae [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2226-2235. |
| [3] | Chen Huiting, Zeng Weiquan, Zhou Jianhong, Wang Jie, Zhuang Congying, Chen Peiyou, Liang Zeqian, Deng Weiming. Tail anchoring technique of vertebroplasty in treatment of osteoporotic vertebral compression fractures with intravertebral cleft: a finite element analysis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2145-2152. |
| [4] | Zeng Xuan, Weng Rui, Ye Shicheng, Tang Jiadong, Mo Ling, Li Wenchao. Two lumbar rotary manipulation techniques in treating lumbar disc herniation: a finite element analysis of biomechanical differences [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2153-2161. |
| [5] | Cheng Qisheng, Julaiti·Maitirouzi, Xiao Yang, Zhang Chenwei, Paerhati·Rexiti. Finite element analysis of novel variable-diameter screws in modified cortical bone trajectory of lumbar vertebrae [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(9): 2162-2171. |
| [6] | Wu Yanting, Li Yu, Liao Jinfeng. Magnesium oxide nanoparticles regulate osteogenesis- and angiogenesis-related gene expressions to promote bone defect healing [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(8): 1885-1895. |
| [7] | Chen Haojie, Wang Dai, Shen Shan. Immune inflammatory microenvironment mechanisms in peri-implantitis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(8): 2054-2062. |
| [8] | Jiang Xinghai, Song Yulin, Li Dejin, Shao Jianmin, Xu Junzhi, Liu Huakai, Wu Yingguo, Shen Yuehui, Feng Sicheng. Vascular endothelial growth factor 165 genes transfected into bone marrow mesenchymal stem cells to construct a vascularized amphiphilic peptide gel module [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(8): 1903-1911. |
| [9] | Hu Xiongke, Liu Shaohua, Tan Qian, Liu Kun, Zhu Guanghui. Shikonin intervention with bone marrow mesenchymal stem cells improves microstructure of femur in aged mice [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1609-1615. |
| [10] | Yuan Xiaoshuang, Yang Xu, Yang Bo, Chen Xiaoxu, Tian Ting, Wang Feiqing, Li Yanju, Liu Yang, Yang Wenxiu. Effect of conditioned medium of diffuse large B-cell lymphoma cells on proliferation and apoptosis of human bone marrow mesenchymal stem cells [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1632-1640. |
| [11] | Li Zhenyu, Zhang Siming, Bai Jiaxiang, Zhu Chen. Osthole improves osteogenic differentiation function of bone marrow mesenchymal stem cells under high-glucose conditions [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1641-1648. |
| [12] | Han Nianrong, Huang Yifei, Akram · Osman, Liu Yanlu, Hu Wei . Programmed cell death receptor-1 suppresses osteogenic differentiation of rat bone marrow mesenchymal stem cells in a high-glucose microenvironment [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1649-1657. |
| [13] | Jin Dongsheng, Zhao Zhanghong, Zhu Ziyin, Zhang Sen, Sun Zuyan, Deng Jiang. Effects of icariin-loaded microsphere-three-dimensional scaffold on osteogenic differentiation of rabbit bone marrow mesenchymal stem cells [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1658-1668. |
| [14] | Zou Yulian, Chen Chaopei, Huang Haixia, Lan Yuyan, Liu Min, Huang Ting. Resveratrol promotes osteogenic differentiation of bone marrow mesenchymal stem cells in an inflammatory microenvironment [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1669-1678. |
| [15] | Liu Anting, Lu Jiangtao, Zhang Wenjie, He Ling, Tang Zongsheng, Chen Xiaoling. Regulation of AMP-activated protein kinase by platelet lysate inhibits cadmium-induced neuronal apoptosis [J]. Chinese Journal of Tissue Engineering Research, 2026, 30(7): 1800-1807. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||