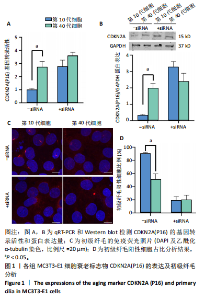
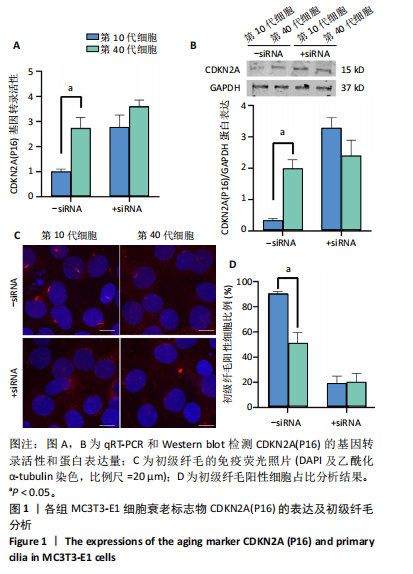
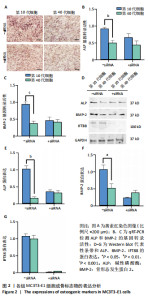
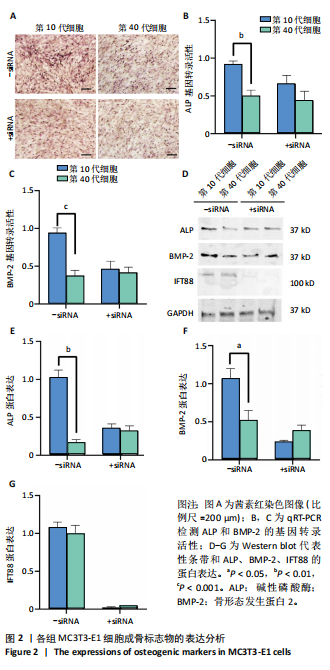
[1] 张元平,崔继秀,裴国献,等.神经化组织工程骨构建的初步观察[J].中华创伤骨科杂志,2005,7(1):60-65.
[2] 秦宇星,任前贵,沈佩锋.组织工程骨技术治疗骨缺损的优越性[J].中国组织工程研究,2020,24(24):3877-3882.
[3] ATASOY-ZEYBEK A, HAWSE GP, NAGELLI CV, et al. Transcriptomic changes during the replicative senescence of human articular chondrocytes. bioRxiv [Preprint]. 2023:2023.11.07.565835.
[4] ALVES-PAIVA RM, DO NASCIMENTO S, DE OLIVEIRA D, et al. Senescence State in Mesenchymal Stem Cells at Low Passages: Implications in Clinical Use. Front Cell Dev Biol. 2022;10:858996.
[5] CUI J, SHIBATA Y, ZHU T, et al. Osteocytes in bone aging: Advances, challenges, and future perspectives. Ageing Res Rev. 2022;77:101608.
[6] PETERSON WJ, TACHIKI KH, YAMAGUCHI DT. Serial passage of MC3T3-E1 cells down-regulates proliferation during osteogenesis in vitro. Cell Prolif. 2004;37(5):325-336.
[7] CHUNG CY, IIDA-KLEIN A, WYATT LE, et al. Serial passage of MC3T3-E1 cells alters osteoblastic function and responsiveness to transforming growth factor-beta1 and bone morphogenetic protein-2. Biochem Biophys Res Commun. 1999;265(1):246-251.
[8] GALLAGE S, GIL J. Primary cilia and senescence: a sensitive issue. Cell Cycle. 2014;13(17):2653-2654.
[9] MA R, KUTCHY NA, CHEN L, et al. Primary cilia and ciliary signaling pathways in aging and age-related brain disorders. Neurobiol Dis. 2022;163:105607.
[10] SILVA DF, CAVADAS C. Primary cilia shape hallmarks of health and aging. Trends Mol Med. 2023;29(7):567-579.
[11] MCMURRAY RJ, WANN AK, THOMPSON CL, et al. Surface topography regulates wnt signaling through control of primary cilia structure in mesenchymal stem cells. Sci Rep. 2013;3:3545.
[12] YAN JL, ZHOU J, MA HP, et al. Pulsed electromagnetic fields promote osteoblast mineralization and maturation needing the existence of primary cilia. Mol Cell Endocrinol. 2015;404:132-140.
[13] VAGHJIANI VG, COCHRANE CR, JAYASEKARA WSN, et al. Ligand-dependent hedgehog signaling maintains an undifferentiated, malignant osteosarcoma phenotype. Oncogene. 2023;42(47):3529-3541.
[14] DI MICCO R, KRIZHANOVSKY V, BAKER D, et al. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat Rev Mol Cell Biol. 2021;22(2):75-95.
[15] DING P, GAO C, GAO Y, et al. Osteocytes regulate senescence of bone and bone marrow. Elife. 2022;11:e81480.
[16] SFEIR JG, DRAKE MT, KHOSLA S, et al. Skeletal Aging. Mayo Clin Proc. 2022;97(6):1194-1208.
[17] 秦宇星,任前贵,沈佩锋,等.组织工程技术治疗骨缺损:应用于临床还有多远?[J].中国组织工程研究,2021,25(29):4703-4708.
[18] 陈裕浩,朱向情,潘兴华.骨髓间充质干细胞的衰老特征及其与治疗的相关性[J].中国组织工程研究,2017,21(17):2746-2752.
[19] TU Z, WANG X, CAI H, et al. The cell senescence regulator p16 is a promising cancer prognostic and immune check-point inhibitor (ICI) therapy biomarker. Aging (Albany NY). 2023;15(6):2136-2157.
[20] SHEN T, GAO K, HU Z, et al. Ciliary Mechanism of Regulating Hedgehog and Wnt/β-Catenin Signaling Modulates Ultraviolet B Irradiation-Induced Photodamage in HaCaT Cells. J Biomed Nanotechnol. 2019; 15(1):196-203.
[21] WANN AK, KNIGHT MM. Primary cilia elongation in response to interleukin-1 mediates the inflammatory response. Cell Mol Life Sci. 2012;69(17):2967-2977.
[22] INGHAM PW. Hedgehog signaling. Curr Top Dev Biol. 2022;149:1-58.
[23] LI B, YAN YP, HE YY, et al. IHH, SHH, and primary cilia mediate epithelial-stromal cross-talk during decidualization in mice. Sci Signal. 2023;16(774):eadd0645.
[24] SHAO YY, WANG L, WELTER JF, et al. Primary cilia modulate Ihh signal transduction in response to hydrostatic loading of growth plate chondrocytes. Bone. 2012;50(1):79-84.
[25] THOMPSON CL, PLANT JC, WANN AK, et al. Chondrocyte expansion is associated with loss of primary cilia and disrupted hedgehog signalling. Eur Cell Mater. 2017;34:128-141.
[26] AL-AZAB M, WANG B, ELKHIDER A, et al. Indian Hedgehog regulates senescence in bone marrow-derived mesenchymal stem cell through modulation of ROS/mTOR/4EBP1, p70S6K1/2 pathway. Aging (Albany NY). 2020;12(7):5693-5715.
[27] HUANG W, HICKSON LJ, EIRIN A, et al. Cellular senescence: the good, the bad and the unknown. Nat Rev Nephrol. 2022;18(10):611-627.
[28] NGUYEN TD, TRUONG ME, REITER JF. The Intimate Connection Between Lipids and Hedgehog Signaling. Front Cell Dev Biol.2022;10:876815.
[29] LV B, ZHANG XO, PAZOUR GJ. Arih2 regulates Hedgehog signaling through smoothened ubiquitylation and ER-associated degradation. J Cell Sci. 2022;135(16):jcs260299.
[30] GREVENSTEIN D, OPPERMANN J, WINTER L, et al. First detection of primary cilia in injured human anterior cruciate ligament: A pilot study with pathophysiological reflections. Pathol Res Pract. 2022;237:154036.
[31] YANG S, ZHENG X. The Immunofluorescence-Based Detection of Hedgehog Pathway Components in Primary Cilia of Cultured Cells. Methods Mol Biol. 2022;2374:89-94.
[32] 马占华,严旭,姜岩,等.初级纤毛/鞭毛转运系统介导力学反应性信号通路促骨髓基质干细胞成骨分化[J].中国组织工程研究, 2024,28(25):3937-3941.
[33] 王小健,张阳,郭丽,等.慢性肾功能不全幼鼠胫骨生长板软骨细胞初级纤毛表达对细胞增殖的影响[J].中华医学杂志,2019, 99(28):2225-2229.
[34] MITRA A, LOSEVA E, PETERMAN EJG. IFT cargo and motors associate sequentially with IFT trains to enter cilia of C. elegans. Nat Commun. 2024;15(1):3456.
[35] TENG YN, CHANG HC, CHAO YY, et al. Etoposide Triggers Cellular Senescence by Inducing Multiple Centrosomes and Primary Cilia in Adrenocortical Tumor Cells. Cells. 2021;10(6):1466.
[36] LeE J, YI S, WON M, et al. Loss-of-function of IFT88 determines metabolic phenotypes in thyroid cancer. Oncogene. 2018;37(32):4455-4474.
|